
Abstract
Salmonella is a leading cause of foodborne illness in the United States. It is a zoonotic pathogen found in many species of food animals, and contamination of foodstuffs by strains of Salmonella found on farms is an important source of human exposure. Here we describe a long-term (2004–2010) study of Salmonella colonization on a typical dairy farm in the Northeastern United States. The fecal shedding prevalence in the herd ranged from 8% to 97%, and greater than 50% of the herd was shedding Salmonella for more than two-thirds of the study period. Salmonella enterica serotype Cerro was first detected in September 2004, after a small and very short-lived outbreak of Salmonella Kentucky. Cerro persisted within the herd for over 3 years, with no clinical signs of salmonellosis in the animals. In the winter of 2006, Kentucky was again detected within the herd, and over a 2-year period, Kentucky gradually supplanted Cerro. Kentucky was the only serotype detected from March 2008 until September 2009, when Cerro was again detected in 15% of the cows on the farm. Since September 2009, Kentucky and Cerro have coexisted within the herd, which continues to harbor these serotypes at high prevalence. Pulsed-field gel electrophoresis (PFGE) could not discern differences between Cerro strains isolated during this study, but it did suggest that the strain of Kentucky that seemed to behave as a commensal in these dairy cows is distinct from the transient strain isolated in 2004. Understanding the dynamics of competition between these two serotypes that seem to behave as commensal colonizers of dairy cows may provide insights into the mechanisms by which Salmonella establishes infection in the lower gut of dairy cows and may lead to the development of measures to prevent or limit Salmonella colonization of dairy cows.
Introduction
We previously described a long-term subclinical outbreak of Salmonella enterica serotype Cerro in a Pennsylvania dairy herd (Van Kessel et al., 2007). This herd was a participant in a longitudinal study of the dynamics of infectious diseases (Pradhan et al., 2009). The outbreak was first detected in September 2004, when Salmonella was isolated from the feces of 44% of the adult cows. None of the infected animals were exhibiting signs of illness during this time. The herd was monitored every 6–8 weeks, and Salmonella-positive cows were identified at each sampling. Although the fecal shedding prevalence dropped to 8% at one sampling, the shedding rate mostly remained above 30%, and at the highest prevalence, 88% of the cows were shedding Salmonella Cerro. This constant shedding led to frequent detection of Salmonella in milk filters and bulk tank milk (Van Kessel et al., 2008). By most measures, this was a well-managed herd with no obvious breeches in biosecurity, manure handling, or hygiene protocols (Pradhan et al., 2009).
We continued to monitor this herd for fecal Salmonella shedding following the publication of the Salmonella Cerro outbreak. We describe here the dynamics of changes in the Salmonella serotype population that infected this herd during a ∼5-year period.
Methods
Farm description
The farm and herd characteristics have been described previously (Van Kessel et al., 2007, 2008). Briefly, the farm maintained 100–110 milking cows housed in a free stall barn. Replacement animals were primarily obtained from within the herd, although two were purchased during the study period. Heifers were moved to an off-site facility at approximately 6 months of age and were returned to the farm prior to first gestation (20–24 months).
Sample collection
All adult cattle (dry cows, lactating cows, and pre-fresh heifers) were sampled at each collection. The sampling interval was 6–12 weeks. Fecal grabs were collected directly from the rectum. Fecal samples were collected from some calves on the day that they were moved to the heifer growing facility and from the heifers when they were moved back to the herd. Milk and in-line milk filters were collected weekly.
Bacterial analysis
Methods for culture and isolation of Salmonella were previously described (Van Kessel et al., 2007, 2008). Briefly, samples were enriched in tetrathionate broth (BD Diagnostics, Sparks, MD) and struck onto XLT4 agar (XLT4 agar base with XLT4 supplement; BD Diagnostics, Sparks, MD) for isolation. Isolated, presumptive Salmonella colonies (five or six randomly chosen isolates per sample) were transferred from XLT4 plates onto XLT4, Brilliant Green, and L-agar (Lennox Broth base with 1.5% agar; Gibco Laboratories, Long Island, NY) and incubated at 37°C for 24 h. Colonies exhibiting the Salmonella phenotype were preserved (−80°C) from the L-agar for future analysis.
As described previously (Van Kessel et al., 2007), from March 2004 to January 2006, serotypes of Salmonella isolates were determined using rep-polymerase chain reaction (PCR). After January 2006, isolates were serogrouped using the PCR method described by Herrea-Leon et al. (2007). Based upon the known history of serotypes on this farm, isolates characterized as serogroup C2 were considered to be Kentucky, whereas those that could not be placed in a serogroup were considered to be Cerro (group K). At least one isolate per positive sample was serogrouped. Random isolates (44 Cerro and 32 Kentucky) were sent to the National Veterinary Services Laboratories (NVSL; Ames, IA) for serotype confirmation, and in all cases the PCR-based serotype designation was confirmed. Additionally, pulsed-field gel electrophoresis (PFGE), as described below, confirmed the serotype designation of 28 Cerro isolates and 43 Kentucky isolates.
A subset of isolates were further characterized by PFGE following the standardized PulseNet Salmonella protocol described by Ribot et al. (2006), with a few modifications. Thiourea (50 μM) was added to the running gel and the electrophoresis buffer (Murase et al., 2004). Run conditions followed the method of Xi et al. (2008). PFGE profiles were analyzed using BioNumerics software (Applied Maths, Austin, TX), with bands assigned manually. Dendrograms were generated from the average of the XbaI and BlnI experiments using Unweighted Pair Group Method with Averages (UPGMA) cluster analysis.
Results
Fecal samples were collected from the milking herd 37 times between March 2004 and December 2010, and cultured for Salmonella. Except for the initial sampling in March 2004, where just one animal was shedding Salmonella Typhimurium, the lowest shedding prevalence was 8% (9/107) in August 2005 (Fig. 1). There was also a low shedding period in December 2008, when salmonellae were isolated from only 14 of 108 cows (13%). The highest prevalence period was between March and September of 2010; nearly all of the animals were shedding in June 2010 when 95 of 98 (96.9%) cows at the farm were culture-positive for Salmonella. Greater than 50% of the herd was shedding Salmonella for more than two-thirds of the study period.

Fecal Salmonella shedding prevalence in a ∼100 cow dairy herd from March 2004 to December 2010. The number of cows in the herd ranged from 98 to 113 during the study.
The producer collected fecal samples from 178 calves immediately before they were transported to the heifer growing facility, and all of these samples were Salmonella-negative. The returning pre-fresh heifers were generally put into the dry cow free stall pen. Fecal samples were collected from 164 of these heifers, and 35 (21.3%) were Salmonella-positive. The first heifers that were identified as Salmonella-positive were returned to the milking herd in November 2005. Serotypes Cerro and Kentucky were both identified amongst the isolates from these animals.
Weekly bulk milk samples were collected, with the exception of a few holiday weeks. In total, 336 milk samples were collected, and Salmonella was isolated from 23 (6.8%). Weekly collection of in-line milk filters began in November 2004. Salmonella was isolated from 183 of the 305 filters collected (60%).
As reported previously, the predominant serotype isolated during the initial Salmonella outbreak from September 2004 to January 2006 was Cerro. Other serotypes (Kentucky, Oranienburg) were occasionally identified during this time, but in less than 0.5% of the samples. As the sampling continued, isolate serotypes were not routinely determined due to the cost. However, when random isolates were characterized in the spring of 2006, although Cerro was still predominant, serotype Kentucky was identified with increasing frequency. Based on previous work with in-line milk filters (Van Kessel et al., 2008), we attempted to use milk filter isolates to determine the serotype distribution in the herd. Depending on availability, the serotype was determined on as many as 10 isolates from each Salmonella-positive filter (Fig. 2). When no obvious relationships were identified between predominant fecal serotypes and milk filter serotypes, it was deemed necessary to serotype more fecal Salmonella isolates to determine the herd serotype profile.
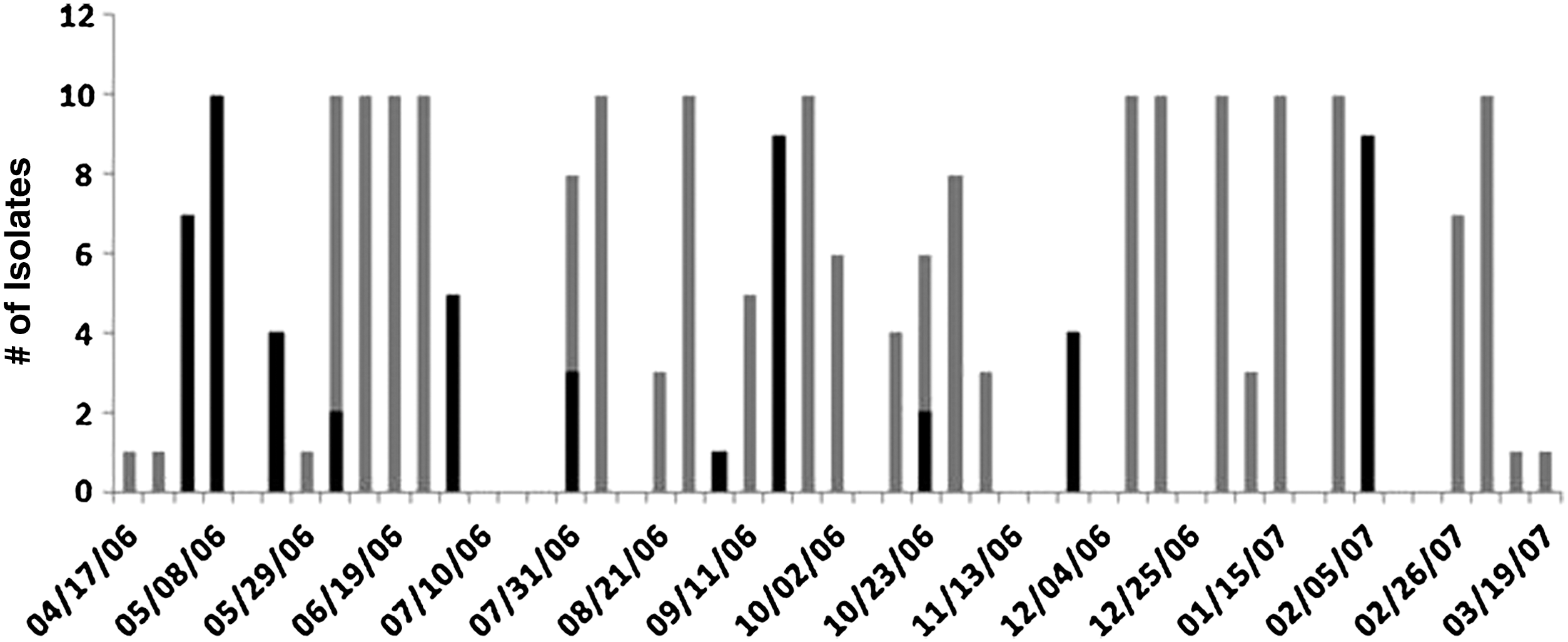
Serotype distribution of Salmonella isolates from in-line milk filters that were collected weekly from the study farm. When available, 10 isolates were serogrouped for each positive sample. When fewer than 10 isolates were available, the serogroup was determined on all of the isolates from that positive sample. Black, Cerro; gray, Kentucky.
When the serotypes of Salmonella isolates were determined for at least one isolate per positive fecal sample, a gradual shift from serotype Cerro to serotype Kentucky was observed, from January 2006 to June 2008 (Fig. 3). Only one cow was shedding Salmonella Cerro in October 2007, and she continued to shed this serotype until she was culled from the herd in June 2008. Kentucky was the only serotype identified in the herd from June 2008 until the sampling in September 2009, when 46% of the herd was shedding Salmonella Kentucky, but 16% of the herd was shedding Salmonella Cerro. Generally, Kentucky remained the dominant serotype in the herd until the last sampling in December 2010.
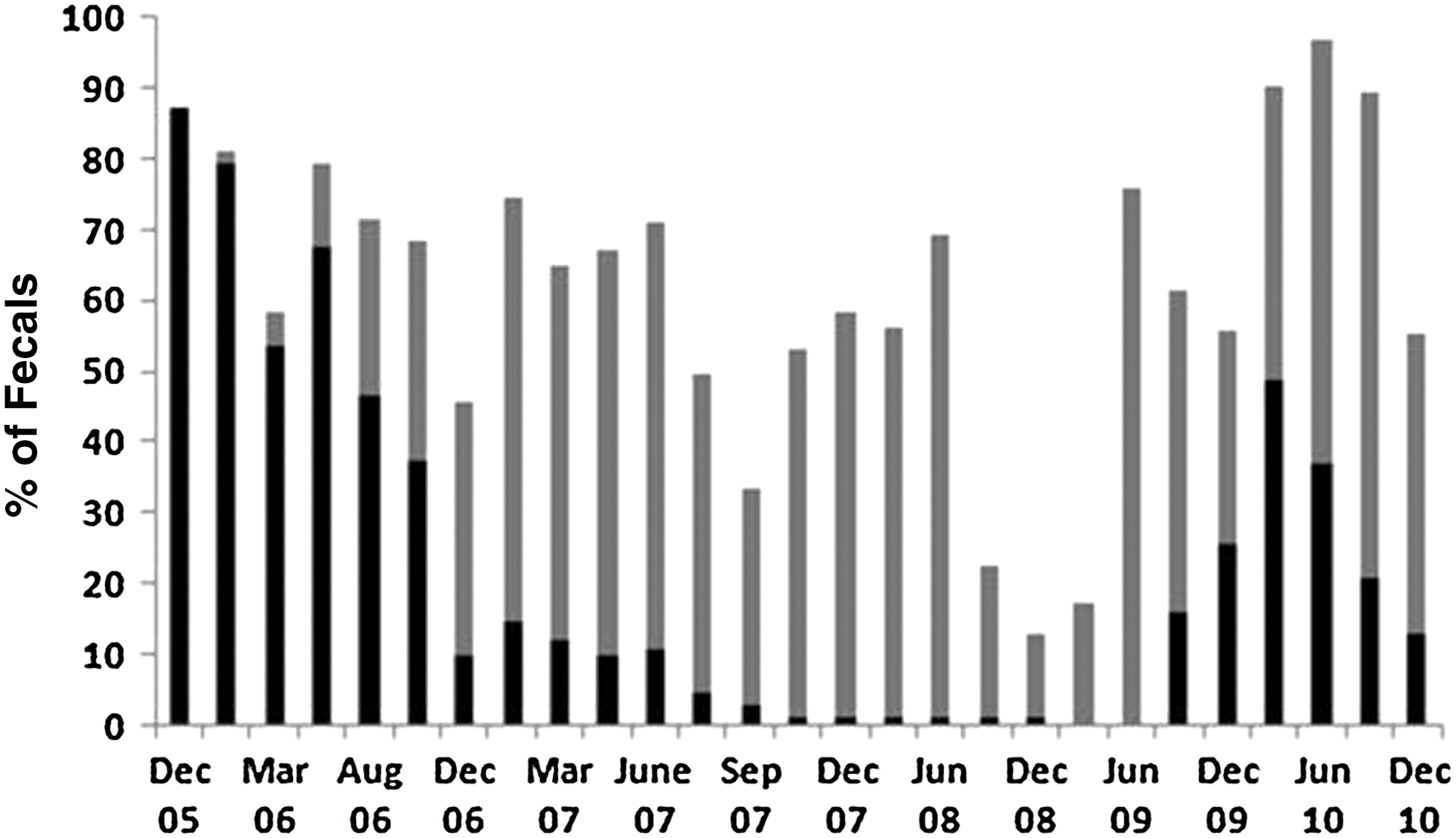
Serotype distribution of fecal Salmonella shedding in a ∼100 cow dairy herd from December 2005 to December 2010. Prior to December 2005, Cerro was always the predominant serotype. Black, Cerro; gray, Kentucky.
Isolates of Salmonella Cerro and Salmonella Kentucky representing the range of time during which each serotype was isolated from the farm were characterized by PFGE. When the restriction digest patterns of the Salmonella Cerro isolates were compared, there was very little variation (>95% similarity), even when the patterns from isolates collected in 2004 were compared with those collected in 2010 (Fig. 4). In contrast, there was somewhat more variation in the restriction digest patterns of the Salmonella Kentucky isolates, and these isolates grouped into two distinct clades (88% similarity between clades). Strains that were isolated in 2004 were grouped into clade K1 (98% within clade similarity), whereas clade K2 (94% within clade similarity) contained isolates from the latter phases of the study when Salmonella Kentucky was more prevalent.

Dendrogram generated from the average of two enzyme (Xba1 and Bln1) pulsed-field gel electrophoresis (PFGE) patterns for representative Salmonella Kentucky and Salmonella Cerro isolates.
Discussion
Dairy cattle are known reservoirs of Salmonella, and numerous studies have identified this pathogen in lactating cows, replacement heifers, and calves, as well as in many locations throughout the farm environment. Based on the National Animal Health Monitoring System (NAHMS) Dairy Surveys conducted in 1996, 2002, and 2007, the fraction of U.S. dairy operations infected with Salmonella has increased (Blau et al., 2005; USDA, 2011). The percentage of positive dairy operations nearly doubled from 1996 when Salmonella was isolated from 21.1% of the herds to 2007 when 39.7% of the herds were identified as Salmonella-positive. While herds selected for fecal sampling in the NAHMS surveys were convenience samples and not sufficient in number to provide true national estimates, these are the only studies that provide an indication of national trends in Salmonella prevalence on U.S. dairy operations. Regional surveys have also been conducted, and although the results showed a wide range of herd prevalence, they also indicated a significant presence of Salmonella in dairy herds (Callaway et al., 2005; Cummings et al., 2009; Fossler et al., 2004; Pangloli et al., 2008; Rodriguez et al., 2006).
Dairy cattle harbor many Salmonella serotypes that are not predominant human pathogens. The fact that some serotypes have not been identified in foodborne outbreaks may be, in some cases, due to lack of exposure. Although some Salmonella serotypes are adapted to specific animal hosts (such as Dublin and cattle), most are not host-specific and can infect multiple species. Multiple serotypes are frequently isolated from a herd and even from a single animal. As with humans, cows infected with Salmonella can exhibit a range of mild to severe symptoms, but non-clinical, asymptomatic shedding, as seen in this study, is also common. There were no obvious signs of illness or adverse affects on production associated with the Salmonella Cerro or the Salmonella Kentucky infections in this herd over the 5+ years of this study.
Salmonella Cerro and Kentucky have been frequently isolated from dairy cows, and based on NAHMS Dairy Surveys, both of these serotypes appear to be increasing in prevalence in U.S. dairy herds (USDA, 2011). Cerro, along with Mbandaka and Typhimurium, were the third most common serotypes isolated from feces and environmental samples in 1996; Kentucky was ranked seventh along with Give, Muenchen, and Senftenberg. In the 2007 survey, Cerro and Kentucky were the most commonly isolated serotypes from feces and environmental samples. Although Cerro was not among the top seven serotypes in the 2002 testing of fecal samples, it was isolated from the bulk milk of two operations (Van Kessel et al., 2004). None of the 2002 Salmonella isolates from bulk milk were serotype Kentucky. Milk filters were sampled in 2007 in addition to bulk milk, and Cerro and Kentucky were the most predominant serotypes isolated (Van Kessel et al., 2011). Cerro was isolated from either a milk or filter sample from 24% of dairy operations that had a positive sample, whereas Kentucky was isolated from 13.7% of the operations that had Salmonella-positive milk and/or filters. Based on these data, Cerro and Kentucky now appear to be very common Salmonella serotypes on U.S. dairy operations.
Sampling group pens and alleyways is generally effective at detecting Salmonella shedding in a herd. Previous work indicated that monitoring milk filters in this herd was also a useful tool for not only detecting the presence of Salmonella shedding in the herd, but also for estimating the number of infected animals (Van Kessel et al., 2008). Therefore, it seemed possible that isolates from the filters would be indicative of the distribution of serotypes in the herd. However, when the serotype profiles of the filter isolates and the concurrent fecal isolates were compared, they were very different. The profile of the milk filter isolates reflects what gets into the milking system, and this is not necessarily representative of what is being shed by the animals. Feces and debris enter the system when there are breeches in the hygiene protocol, and therefore the serotype distribution in the filter can be influenced by the shedding status of the animal being milked at the time of a breech.
Although infection with Salmonella Cerro did not appear to have clinical impact on the cows in the herd described here, Cummings et al. (2010) suggested that Cerro is potentially an emerging pathogen of dairy cattle. In their study of dairy cattle in New York, Cummings et al. (2010) identified Salmonella Cerro in 20 of 44 Salmonella-positive herds. Salmonellosis-like symptoms were reported in some, but not all of these herds. Cummings et al. (2010) also cultured feces from animals that were suspected of having salmonellosis, and Cerro was identified as the main serotype in 59% of these samples. Isolates of Cerro from clinical bovine sources have also been identified by NVSL (CDC, 2008). It is unclear if Salmonella Cerro infection caused the disease in these cases, but in the Cummings et al. (2010) survey, Cerro was identified more frequently from clinical fecal specimens than from feces of healthy animals. It is also unclear why the prevalence of Salmonella Cerro appears to be increasing in dairy cattle; perhaps Cerro is gradually adapting to the animals and the dairy environment, or it is more simply a measure of increasing exposure or pressure. When isolates of Cerro from the study herd were compared by PFGE, there was very little distinction, either when isolates represented the 5+ year span of the study or when they were compared across animals. Cerro isolates collected from bulk milk and milk filters in the NAHMS Dairy studies (Van Kessel et al., 2004, 2011) were from multiple states, and no distinction could be made based on PFGE. Other studies have shown similar clonality within Cerro from diverse geographical sources (Hoelzer et al., 2011).
The serotype Kentucky is more commonly associated with human illness than Cerro but is still relatively rare (CDC, 2008). Isolates from non-human sources come primarily from cattle and poultry, and occasionally from swine (CDC, 2008). As with Salmonella Cerro, Salmonella Kentucky appears to have variable impact on cattle, and has been identified in asymptomatic cows and in cows exhibiting salmonellosis-like symptoms. Kentucky is a more genotypically diverse serotype than Cerro, and PFGE analysis can be useful to discriminate between isolates (Xi et al., 2008). When PFGE analysis was conducted on Kentucky isolates from 2004 and from the later phase of the study where Kentucky established a presence in the herd, two major clades (Fig. 4) were observed. Isolates from 2004 (K1) were distinct from isolates obtained when Kentucky began to establish a presence in the herd (K2). It is possible that this observed diversity is related to the difference in ability of the strains to establish in the herd, but more investigation is needed.
Feed components and total mixed rations were tested throughout the study and were not an apparent source of Salmonella on this farm (data not shown). The earliest isolates of Salmonella Kentucky that group into clade K2 came from a water trough at the heifer-raising facility and from a returning pre-fresh heifer fecal collected in November 2005. This suggests that the practice of off-site heifer rearing was the breech in biosecurity that led to the Salmonella Kentucky infection in this herd. Off-site heifer-raising operations have previously been identified as potential sources of disease for milking herds, particularly when the operation services multiple herds (Hegde et al., 2005).
The unique aspect of this study is frequent, in-depth sampling of a single farm over multiple years. This longitudinal sampling regime revealed the dynamics of asymptomatic Salmonella infection in an endemically infected dairy herd from the onset of infection through two serotype shifts. The transition from Cerro to Kentucky was very gradual and was followed by an extended period where Kentucky predominated. A second serotype shift led to the re-establishment of Cerro in the herd. These population shifts are evidence of competition between Salmonella strains within a dairy herd. Even though other serotypes were isolated at various times throughout the study, they were never able to establish residency within the herd. It is possible that the endemic infection with Kentucky and Cerro blocked infection by other serotypes. Alternatively, Kentucky and Cerro may be more prevalent in the environment surrounding the dairy and the external pressure of these serotypes was greater.
It is likely that these long-term infections and population shifts are found on many dairy farms but they are usually only identified when there is an impact on animal health and production. Understanding the factors that lead to the initial infection and subsequent competition between commensal strains such as Cerro and Kentucky may help identify factors involved in the colonization of dairy cows by salmonellae. This in turn may lead to the development of strategies for preventing or eliminating endemic Salmonella infections. More efforts are needed to identify mitigation strategies that are effective to reduce the prevalence of all salmonellae on dairy farms. Pre-harvest control of zoonotic pathogens is critical for lowering or minimizing the potential exposure of the public to contamination by bacterial pathogens in the foods that they eat.
Footnotes
Acknowledgments
We gratefully acknowledge the assistance of the support staff at the Department of Veterinary and Biomedical Sciences, Pennsylvania State University, and at the Environmental and Microbial Food Safety ARS Laboratory in Beltsville. We also appreciate the tremendous cooperation of the farm owner.
Disclosure Statement
No competing financial interests exist.