
Abstract
The objective of this study was to characterize the epidemiology of Salmonella carried by dairy cows culled from herds in the Texas High Plains. Feces were collected from a convenience sample of 706 animals culled from nine dairy farms. In addition, individually paired fecal and hide samples were collected from 70 healthy milking cows on three of the dairies. Samples were cultured for Salmonella using routine methods; isolates were serotyped and subjected to a panel of antimicrobial drugs to determine susceptibility. Salmonella was recovered from 32.6% of culled cows. Whole-herd use of a vaccine containing siderophore receptors and porin proteins was associated (p=0.05) with reduced Salmonella prevalence in that the prevalence among herds that practiced whole-herd vaccination was 8.0% compared to 36.8% among herds that did not use this vaccine. The majority (88.6%) of isolates were pansusceptible or resistant to one drug. Of the 3.1% of isolates resistant to more than four drugs, all were Salmonella Newport and were recovered from one dairy. Various serotypes were recovered from individual fecal and hide samples. Salmonella Montevideo was recovered more frequently (p<0.01) from hide samples, whereas Salmonella Cerro was recovered more frequently (p<0.01) from feces. Salmonella was recovered from at least one cow on all dairies. While our study was not a priori designed to address herd-level factors, we found evidence that the whole-herd use of a siderophore receptor and porin protein-containing vaccine might be a useful aid in the control of Salmonella in groups of cattle. As this is a nonrandomized evaluation of an intervention, other herd-level factors that may be correlated with vaccine use, such as biosecurity, might have been responsible for the observed association.
Introduction
While not the most commonly attributed source of infection, beef is nevertheless an important vehicle for exposure (McLaughlin et al., 2006; Guo et al., 2011; Schneider et al., 2011). Despite the complex, segmented nature of beef production in the United States, which makes animal-source attribution difficult (Schneider et al., 2011), several outbreaks of salmonellosis, including instances of disease caused by MDR variants, have been linked to beef harvested from culled dairy cows (McLaughlin et al., 2006; Schneider et al., 2011). On dairy farms, cows may be culled from the milking herd for various reasons such as low productivity (Ahlman et al., 2011). These cull (or market) cows are routinely sold for slaughter for human consumption and constitute a substantial proportion of beef production in the United States. Characteristics of the carcasses from cull cows mean that much of the beef is fabricated into components that will be incorporated into ground-beef products (Stelzleni et al., 2007). Furthermore, lean beef trimmings from cull cows are often blended with high-fat content beef trimmings harvested from animals finished in feedlots to facilitate a consistent supply of ground beef that meets certain purchase specifications. As a consequence, beef from culled dairy cows may be broadly incorporated into ground beef products across the United States.
Supported by empirical evidence (Varma et al., 2006; Withee et al., 2009; Williams et al., 2010; Dodd et al., 2011)), many have associated slaughter of pathogen-harboring cattle with adverse public-health events (e.g., Escherichia coli O157-induced foodborne disease). If so, control at various points of the production-consumption continuum could reduce human exposure to various pathogens. To date, most efforts to control foodborne pathogens have occurred within slaughter establishments. It is possible, however, that efficacious and practical pre-harvest strategies may further contribute to meaningful control of pathogens, such as Salmonella. In prior research (Loneragan and Brashears, 2005; Kunze et al., 2008), Salmonella was commonly recovered from cattle in the southern High Plains of Texas, and when identified, MDR Salmonella were more likely to be recovered from culled dairy cows than from healthy feedlot animals (Kunze et al., 2008). It was uncertain, however, if the Salmonella was clustered within a few dairies or was broadly disseminated across many herds. Such knowledge is important because it informs the design and implementation of pre-harvest strategies to control Salmonella in that, if Salmonella is clustered within a few herds, strategies might be targeted and herd-specific, but if Salmonella is broadly disseminated, strategies would need to be widely adopted. The objective of this study, therefore, was to characterize the epidemiology of Salmonella carried by dairy cows culled from herds in the Texas High Plains.
Methods
A convenience sample of nine dairies located within a 50-km radius of Dalhart, Texas, was enrolled; criteria for enrollment were a willingness to participate and that most culled cows were sold at a regional auction market. Dairies varied in size from approximately 500–5000 lactating cows, with an average size of approximately 2300 cows. Cow-level enrollment was a convenience sample based on availability of cull cows.
Cow-level fecal samples were collected over 21 days from early June through September. In general, cattle were transported to a regional auction market, and approximately 50–100 g of feces were collected per rectum during pre-sale inspection. Dairy of origin was determined using transportation receipts. On instances when cows were marketed directly to an abattoir from the dairy, thereby bypassing the auction market, samples were collected at the dairy. In addition to the aforementioned samples, paired fecal samples and hide swabs, 1,000 cm2 over the dorsal midline, were collected from a convenience sample of 70 clinically health milking cows on three dairies to explore serotype diversity within animals. Swabs for hide sampling were hydrated in 25 mL of Butterfield's broth; prior to sample collection, free broth was expunged from the swab, the area of hide was swabbed, and then the swab was placed back in the broth.
For feces collected from cull cows, 1 g was inoculated into 9 mL of tetrathionate (TT) broth and 1 g of feces was inoculated into 9 mL of buffered peptone water (BPW). One milliliter of this was then inoculated into 9 mL of Rappaport-Vassiliadis (RV) broth. Tetrathionate and RV broth were incubated at 42°C for 24 h, and the original sample was stored at 4°C. After inoculation, broth (TT and RV) was streaked for isolation onto xylose-lysine-tergitol 4 (XLT4) and hektoen enteric (HE) agar plates, and incubated for 24 h at 37°C; a presumptive result was reported based on the presence of morphologically typical isolates (yellow or red with black centers). Up to three morphologically typical colonies per sample were streaked onto tryptic soy agar (TSA) plates for further characterization.
Salmonella-positive samples were subjected to a 3×5 most-probable number (MPN) technique to estimate Salmonella concentration. One gram of feces was inoculated into 9 mL of BPW, and 1 mL of this was placed in three 9-mL tubes of RV (10−1 MPN tubes); then serial dilutions were performed to obtain 10−2 to 10−5 tubes and incubated at 37°C for 18 h. Tubes were streaked onto XLT4 plates, incubated as above, and then inspected for morphologically typical colonies. A freely available MPN calculator was used to calculate MPN per gram of feces (
Feces collected from clinically healthy milking cows were divided into 20 one-gram aliquots per cow. Ten of these aliquots were inoculated into 10 tubes each containing 9 mL of TT broth, and the remaining 10 aliquots were inoculated into 10 tubes each containing 9 mL of RV broth. For hide samples, 20 one-milliliter aliquots of Butterfield's broth were inoculated into 10 tubes containing 9 mL of TT broth and 10 tubes containing 9 mL of RV broth. Tubes were incubated for 24 h at 42°C, and then streaked for isolation onto XLT4 and HE agar plates and incubated as above. Up to 10 morphologically typical colonies from each sample were streaked onto TSA for further characterization.
All isolates were transferred to 10 mL of brain-heart infusion broth and incubated at 37°C for 24 h. Broth was streaked for isolation onto TSA plates then incubated at 37°C for 24 h. Three to five well-isolated colonies were transferred into 4 mL of sterile, de-ionized water and adjusted to a 0.5 McFarland standard. Ten microliters of this was then transferred into 10 mL of Mueller-Hinton broth and used to inoculate the 96-well micro-broth dilution plates (Sensititre, catalog number CMV1AGNF; TREK Diagnostic Systems, Cleveland, OH; Table 1). At least two plates from each lot were evaluated for quality control with E. coli ATCC 23742. Minimum inhibitory concentration (MIC) was reported as the lowest concentration that inhibited growth; if the isolate grew at the greatest concentration, the MIC was arbitrarily set at double that concentration. Isolates were classified as susceptible or resistant based on available breakpoints (CDC, 2006; CLSI, 2006; 2010). All isolates were serotyped (Pennsylvania Animal Diagnostic Laboratory System).
Format of plates employs twofold dilutions from greatest to lowest concentrations included.
Sensititre (catalog number CMV1AGNF; TREK Diagnostic Systems, Cleveland, OH).
Salmonella prevalence and concentration were analyzed using logistic and linear mixed models methods, respectively (SAS version 9.2; The SAS Institute, Cary, NC). Initial crude (i.e., unadjusted) estimates were generated. Where appropriate, dairy was included in the models as a random variable to control for within-dairy clustering and model-adjusted estimates were generated. A Kappa statistic was calculated to estimate the agreement between paired hide and fecal samples. Various independent variables were evaluated for significance using an α level of 0.10.
Results
Seven-hundred and six samples were collected from cull cows across the months of June (n=117), July (n=199), August (n=223), and September (n=167). Samples per dairy varied from 10 to 165 and averaged 78.4 (median=47; Table 2). Salmonella was recovered from 32.6% (n=230; 95% confidence limits [CL]=29.2, 36.1%) of samples. Controlling for dairy-level clustering, prevalence varied across month of sample collection (p<0.01). Model-adjusted Salmonella prevalence in June, July, August, and September were 17.6% (95% CL=6.9, 38.2), 31.2% (14.4, 56.5), 37.5% (17.9, 62.3), and 34.1% (15.7, 59.1), respectively. Prevalence among samples collected in June was less than that observed in other months (p<0.01 for all comparisons); no variation was detected among July, August, and September (p>0.24 for all comparisons). After accounting for dairy-level clustering, model-adjusted Salmonella prevalence (30.6%) was similar to the crude estimate, but the confidence interval around this estimate was much wider, which was indicative of within-herd clustering (95% CL=12.1, 58.5%).
Dairies that practiced whole-herd vaccination with a product containing siderophore receptors and porin proteins.
Three dairies reported practicing whole-herd vaccination with a commercially available Salmonella vaccine (Salmonella Newport Bacterial Extract; Pfizer Animal Health, Madison, NJ) whereas the remaining six reported no use of this vaccine. Prevalence of Salmonella among herds that practiced whole-herd vaccination (8.0% [95% CL=1.3, 35.6%]) was less (p=0.05; relative risk=0.22) than the prevalence observed on herds that did not practice vaccination (36.8% [15.7, 64.5]).
Fecal concentration of Salmonella varied from −0.5 to 7.0 log10 MPN per gram of feces (Fig. 1) and was not associated with vaccine status. Controlling for dairy, log10-transformed concentration varied across month of sample collection (p=0.02) and increased linearly over time (p=0.03), in that in June, July, August, and September, concentration was 0.61 (95%CL=−0.09, 1.3), 0.61 (−0.03, 1.1), 0.90 (0.41, 1.4), and 1.4 (0.91, 1.9) log10 MPN per gram of feces, respectively. At the herd level, concentration did not appear to be a function of prevalence (p=0.73).
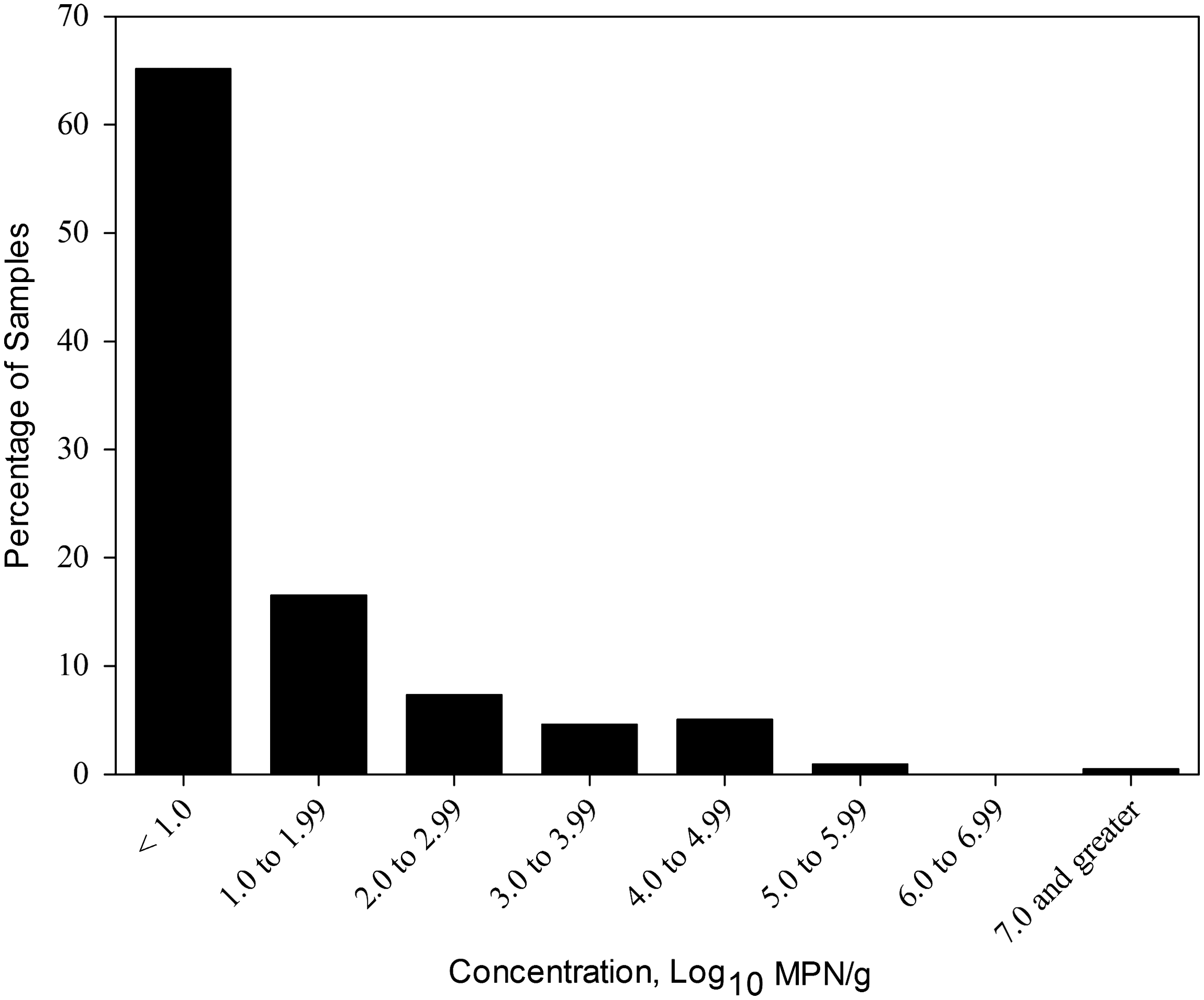
Percentage of samples from which Salmonella was recovered based on estimate of Salmonella concentration reported as log10 MPN/g of feces.
One, two, and three isolates were recovered from 4, 12, and 214 culture-positive samples, respectively, yielding 669 Salmonella isolates for further characterization. In instances where two isolates were recovered per sample, two distinct serotypes were recovered twice (i.e., on 10 occasions, the two isolates recovered were of the same serotype). When three isolates were recovered per sample, two distinct serotypes were identified on 46 occasions and three distinct serotypes were identified seven times. Thus, 292 unique sample-serotype combinations were observed.
Fifteen serotypes were identified (Table 3), and up to 12 serotypes were identified within a dairy. In general, as the prevalence increased, the serotype diversity also increased. The most common number of drugs to which distinct serotype-sample combinations (n=292) was resistant was one drug (56.8%; n=166), followed by pansusceptible (34.9%; 102). Among those isolates resistant to one drug, the most common phenotype was resistance to sulfisoxazole (94.7%, n=157), followed by streptomycin (1.8%; n=3), then tetracycline, cefoxitin, and trimethoprim/sulfamethoxazole (1.2%; n=2 each; Table 4). Resistance to four or more drugs was observed in 4.8% (n=14) of the 292 unique sample-serotype isolates. Of these, the majority (66.7%; n=10) were coresistant to ampicillin, chloramphenicol, streptomycin, sulfisoxazole, and tetracycline (i.e., ACSSuT phenotype), and nine of these were also co-resistant to ceftiofur and potentiated β-lactams (i.e., MDR-AmpC phenotype). All of the 11 isolates that were resistant to more than four drugs were Salmonella Newport and originated on one dairy. Salmonella Newport was recovered from cattle from two other dairies, and these isolates were either pansusceptible or singly resistant to sulfisoxazole.
ACSSuT, phenotypic co-resistance to ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline; MDR-AmpC, phenotypic co-resistance to ACSSuT, a third-generation cephalosporin, and a potentiated β-lactam.
N/A, not applicable in that all isolates in this category were pansusceptible; A, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulfisoxazole; T, tetracycline, Cef, ceftiofur; Fox, cefoxitin; Axo, ceftriaxone; Aug, amoxicillin and clavulonic acid; Tsu, trimethoprim and sulfamethoxazole.
Paired-fecal and hide samples were collected from 70 healthy cows in-milk on three dairies. Of these, Salmonella was recovered from either the feces or hides of 41.4% (n=29) of cows; Salmonella was recovered from 30.0% (n=21) of feces and 34.3% (n=24) of hide samples. There was substantial agreement (beyond chance agreement) between animal-level hide and fecal status (κ=0.58; 95% CL=0.37, 0.78). Multiple serotypes were recovered from 26 of the 40 positive samples in that one serotype was recovered from 13 samples, two serotypes from seven samples, three serotypes from 13 samples, and four serotypes from six samples. Salmonella Montevideo was more commonly recovered from the hide than from feces (p<0.01; Table 5) whereas Salmonella Cerro was more commonly recovered from feces than from hides (p<0.01).
n=number of samples from which at least one isolate was recovered.
Discussion
Salmonella was routinely recovered from cows culled from dairies in the Texas High Plains. We observed some evidence that prevalence increased over time from June and remained elevated through the course of the study; this seasonality is consistent with prior observations in which prevalence during the summer and autumn was greater than that observed during the winter and spring (Kunze et al., 2008). Further, as the study progressed, the concentration of Salmonella increased.
Salmonella was recovered from at least one culled cow from each of the dairies enrolled in our study. The food-safety consequence of this depends on the perspective of what constitutes risk and may vary from one stakeholder to another. For example, if all Salmonella are viewed as a public-health risk or if during the manufacture of ground-beef products any Salmonella may result in failure to meet microbial criteria, then the ubiquitous nature of Salmonella in cull dairy cows may be problematic, and pre-harvest control strategies would necessarily be broadly adopted. In our study, however, the vast majority of the isolates were broadly susceptible to antimicrobial drugs and were of serotypes uncommonly associated with human illness. Consequently, if risk is more narrowly defined (e.g., Salmonella Newport MDR-AmpC), then a more targeted approach to pre-harvest control might be developed.
A need exists to better characterize factors that contribute to Salmonella shedding among cull dairy cows. An ostensibly logical approach would be to explore cow-level factors to discover causes of cases (i.e., factors associated with shedding in individual cows); it is likely that these cow-level factors may be shared among herds. It is important to note, however, that all dairies were younger than 5 years old, and shared many management and health protocols. Furthermore, a salient observation from the study described herein was the tremendous variation in prevalence from dairy to dairy, in that prevalence on some dairies remained consistently and substantially below the average, whereas prevalence on other dairies remained consistently and substantially above the average. Unmeasured herd-level characteristics were, therefore, important determinants of prevalence. An exploration of cow-level factors may consequently miss herd-level determinants of prevalence (Rose, 1985), yet these herd-level factors account for far more variation in prevalence than cow-level factors. Furthermore, because prevalence is a function of both incidence and duration of infection, it is possible these herd-level factors may influence either (or both) to produce the observed variation in prevalence. Potential control strategies, therefore, may be developed based on an understanding of herd-level factors that can reduce the rate of new infections (incidence) or the duration of infection.
Herd-level use of a vaccine containing purified siderophore receptors and porin proteins (SRP) was associated with a 78% reduction in Salmonella prevalence. In a prior evaluation of this vaccine within a single 1,200-cow dairy (Hermesch et al., 2008), an association with Salmonella shedding was not detected, yet it should be noted that the only non-vaccinated cattle were the 90 control animals; consequently, the remaining herd, approximately 1,110 animals, were vaccinated. If this vaccine does induce some degree of protective immunity, it is possible, therefore, that the failure to detect an association in the prior study was due to herd immunity (i.e., herd immunity biased the effect measure toward the null). In the study described herein, we a posteriori explored herd-level vaccine use, and as such, our study was not specifically designed evaluate such an association. Some caution should be used in interpreting the association between vaccine use and Salmonella prevalence because it is possible that unmeasured confounding variables, such as other biosecurity controls implemented in conjunction with the vaccine, may have biased the effect measure. However, our findings warrant further evaluation of this vaccine as an aid in the control of Salmonella in dairy cattle. Further, if herd immunity biases the effect measure toward the null (Peterson et al., 2007), a preferred study design may be one based on herd-level use by allocating dairies to exposed or non-exposed cohorts (i.e., cluster randomized trial), or alternatively, enrollment could be based on existing whole-herd vaccine administration or not (i.e., observational cohort study). If so, investigators should weigh the likelihood and extent of bias arising from herd immunity (i.e., within-herd evaluation) against bias from confounding variables (i.e., herd-level evaluation).
The number of drugs to which Salmonella isolates were resistant displayed a bimodal distribution in that isolates were generally broadly susceptible or broadly resistant and is consistent with findings reported elsewhere for Enterobacteriaceae (Lowrance et al., 2007; Kunze et al., 2008; Platt et al., 2008). While uncommon, the MDR-AmpC phenotype was identified from 3.1% of the 292 unique serotype-sample isolates, yet all of these isolates all came from one herd and all were Salmonella Newport. This serotype, particularly the broadly resistant variant, is one of the most commonly reported causes of clinical salmonellosis in both animals and people (CDC, 2011). The prevalence of all Salmonella on the single herd in which Salmonella Newport MDR-AmpC was identified was 50.0%; of all unique serotype-sample combinations identified on this herd, broadly resistant Salmonella Newport accounted for 39.3% of observations and accounted for all Salmonella Newport on this herd (which did not use the Salmonella SRP vaccine). Seven other serotypes were identified among the remaining 60.7% of isolates. On this operation, therefore, Salmonella Newport MDR-AmpC was the dominant serotype.
Interestingly, we recovered multiple serotypes from both fecal and hide samples collected from healthy cows, in that up to three and four serotypes were recovered from feces and hides, respectively. Not surprisingly, the within-animal serotype distribution reflected the broader serotype diversity observed on the dairy at the time of sample collection. Somewhat unexpectedly, however, we identified a number of serotypes that were differentially recovered from the sample matrices. For example, Salmonella Montevideo was approximately seven times more likely to be recovered from the hides of cattle than their feces. Conversely, Salmonella Cerro was almost twice as likely to be recovered from feces than from hides. The reason for this disparity is uncertain and may reflect subtle physiological differences between serotypes in their ability to survive and persist in various niches. If so, it may inform our understanding of differences in the environmental versus host predilection of various serotypes. Conversely, this finding may be a function of our choice of microbiological methods and their ability to recover various serotypes from different matrices.
Footnotes
Acknowledgment
Dr. Loneragan has provided scientific advice to Pfizer Animal Health and Epitopix, LLC, and has on occasion billed for this service. Dr. Loneragan has also received honoraria for service on advisory boards and presentations.
Disclosure Statement
This research was funded by the Beef Checkoff and National Research Initiative of the USDA Cooperative State Research, Education and Extension Service (grant 2004-35212-14864).