
Abstract
This study aimed to determine the mycological profile of the retail date fruits distributed in different markets at Taif, Saudi Arabia. The presence of aflatoxins and ochratoxin A was also measured. Twenty-two fungal species belonging to 12 genera were isolated from 50 different date samples. Aspergillus flavus, A. niger, Penicillium chrysogenum, and Rhizopus stolonifer were the most prevalent species among isolated fungi. Eighty isolates of A. flavus and 36 of A. niger were detected for their aflatoxins and ochratoxin production potentials using thin layer chromatography. Toxicity test using Artimia larvae indicated that seven out of 18 A. flavus isolates had aflatoxins potentials, while nine out of 36 isolates of A. niger were ochratoxigenic. The quadruplex polymerase chain reaction using specific primers demonstrated the presence of four genes: nor A, ver 1, omt A, and avf A in seven A. flavus toxigenic isolates. Nine A. niger toxigenic isolates showed positive results for the presence of the PKS gene. In conclusion, the present study highlighted the potential hazards of mycotoxins on human health from consuming raw dates. Rapid molecular detection methods described here might help the food authorities to assure the safety of raw dates distributed in local markets.
Introduction
Fresh fruits are prone to fungal contamination in the field; during harvest, transport, and marketing; and with the consumer. Fungi play a substantial role in spoilage of fruits and vegetables, because of their pathogenicity to the harvested products. Fungi (Aspergillus, Alternaria, and Penicillium spp.) may grow on high-moisture dates, especially when harvested following rain or high humidity periods (Kader and Hussein, 2009). Mycological profiles for date palm fruits were studied previously in several places around the world, including Saudi Arabia (Salik et al., 1999; Gherbawy, 2001; Ragab et al., 2001; Shenasi et al., 2002; Aba Alkhail et al., 2004; Alghalibi and Shater, 2004; Iamanaka et al., 2005; Al-Sheikh, 2009; Bokhary, 2010).
During the various stages of pathogenesis, however, some of these fungi may generate different mycotoxins—secondary metabolites that are toxic to humans and animals who consume the products. Aflatoxins are secondary metabolites produced by aflatoxigenic fungi Aspergillus flavus Link and Aspergillus parasiticus Speare (Roehuck and Maxuitenko, 1994). Several researchers documented aflatoxin production in date palm fruits (Mahjoub et al., 1989; Ahmed and Robinson, 1997; Abdel-Sater and Saber,1999; Ragab et al., 2001; Shenasi et al., 2002; Alghalibi and Shater, 2004). Ochratoxin A (OTA) is a nephrotoxic mycotoxin naturally found in a wide range of food commodities throughout the world. In addition to the other toxic effects, OTA is carcinogenic, teratogenic, genotoxic, and immunosuppressive (Petzinger and Weidenbach, 2002). Two Aspergillus sections are known to produce OTA: the section Circumdati (also called the Aspergillus ochraceus group) and the section Nigri (Aspergillus carbonarius and Aspergillus niger) (Varga et al., 1996; Heenan et al., 1998). Among the species of the section Nigri, A. carbonarius shows high ochratoxigenic potential, with most isolates having the ability to produce OTA in culture (Heenan et al., 1998). Sartori et al. (2010) reviewed and discussed the detection of microorganisms capable of producing OTA, prior to ochratoxin production and accumulation. However, very limited information is available in detecting ochratoxin in date fruit (Abdel-Sater and Saber, 1999; Ferracin et al., 2009).
Polymerase chain reaction (PCR)–based methods are considered a good alternative for rapid diagnosis of fungi because of their high specificity and sensitivity (Schmidt et al., 2003), especially when multi-copy sequences are used to develop species-specific primers (Bluhm et al., 2002). The biosynthetic pathway for aflatoxin production by A. flavus has been deciphered, and genes in the aflatoxin biosynthetic pathway have been identified (Payne et al., 1993; Trail et al., 1995; Yu et al., 1995, 2004). The AflR gene plays an important role in the aflatoxin biosynthetic pathway by regulating the activity of other structural genes such as omt-A, ver-1, and nor-1 (Woloshuk et al., 1994; Chang et al., 1999). In particular, aflR gene integrity has been considered a good candidate for diagnostic purposes in discriminating between aflatoxins producer and nonproducer strains of A. flavus (Chang et al., 1995). Reports on the detection of aflatoxigenic fungi using PCR are rather limited (Shapira et al., 1996; Farber et al., 1997; Gashgari et al., 2010, 2011). The Aspergillus niger group is well known as an ochratoxin producer, and several studies have been conducted to explore this group of fungi. Patino et al. (2005) developed two PCR assays to detect A. carbonarius and A. ochraceus, two main sources of OTA contamination, particularly to grapes, coffee, and derivatives in warm climates. Selma et al. (2008) developed a multiplex real- time PCR system for the simultaneous detection of A. carbonarius and A. niger aggregate species. They designed a primer pair and a probe specific for the A. niger aggregate species derived from a conserved region in the acyltransferase domain sequences of PKS genes after comparison with different Aspergillus and Penicillium species. Their method allowed the detection of all the species included in the aggregate, without differentiating between ochratoxigenic and nonochratoxigenic strains. Castellá and Cabañes (2011) designed a primer pair from a PKS gene described in A. niger CBS 513.88 genome that has a strong similarity to the PKS gene of A. ochraceus involved in OTA biosynthesis. This primer pair produced a fragment of 951 bp specific for ochratoxigenic strains of the A. niger aggregate.
The present study aimed to (i) determine the occurrence and load of fungi (the important foodborne pathogens) in date fruits offered for sale to consumers in retail stores at the Taif region in Saudi Arabia; (ii) analyze specifically the occurrence of the Aspergillus niger group and the Aspergillus flavus group in the collected samples; and (iii) investigate the occurrence of OTA and aflatoxins, a public health concern, using chromatographic and molecular techniques.
Methods
Mycobiota determination
Fifty replicated samples of date fruits were collected from different local supermarkets in the Taif region. Fungi were isolated using the dilution plate method as described by Johnson and Curl (1972). Twenty-five grams of each dried fruit sample were comminuted for 2 min in 250 mL of 0.12% sterile plain agar. Further dilutions were made, and 1 mL of an appropriate final dilution was placed on 1% glucose–Czapek's agar plates. The plates were incubated at 28°C for 10 days, and fungal colonies growing on these plates were purified and identified by classical taxonomic methods (Pitt and Hocking, 1997). From media plates, only colonies belonging to A. niger and A. flavus groups were transferred to slants to ensure precise counting and for identification at the species level (Klich and Pitt, 1988). These isolates were preserved at −80°C for further studies.
Mycotoxin detection using thin layer chromatography
Aflatoxins
The isolates identified as Aspergillus species were tested for the production of aflatoxins based on the thin layer chromatography (TLC) method (Rahimi et al., 2007).
OTA
Each isolate from black aspergilli was grown as stationary cultivation and incubated at 28°C for 15 days. Liquid medium of the following composition was used: sucrose, 30 g; peptone, 10 g; NaNO3, 2 g; K2HPO4, 1 g; yeast extract, 1 g; KCL, 0.5 g; MgSO4. 7H2O, 0.5 g; FeSO4. 7H2O, 0.01 g; 1 L of distilled water. The cultivation was made in 250-mL Erlenmeyer flasks. The content of each flask (medium + mycelium) was homogenized for 3 min in a high-speed blender (16,000 rpm) with 75 mL of chloroform. The extraction procedure was repeated three times. The combined chloroform extracts were washed with equal volumes of distilled water, dried over anhydrous sodium sulphate, filtered and then concentrated under a vacuum or a stream of nitrogen to near dryness (El-Kady and Moubasher, 1982). OTA was detected in the sample extracts after TLC on 0.3-mm layers of Adsorbosil 5 silica gel, using tolueneethyl acetate–90% formic acid (6:3:1 vIvIv) for development. Each extract formed a green fluorescent spot under ultraviolet light with the same Rf (0.55) as authentic OTA. The fluorescence turned blue on treatment with ammonia (Nesheim, 1976).
Molecular detection of mycotoxin producing genes
The isolation of DNA from mycelia was performed according to the method described by Farber et al. (1997). Specific primers used in the current study are shown in Table 1. For aflatoxins, PCR assays were performed in 25 μL of a reaction mixture that contained the following: 2.5 μL buffer, 1.5 μL MgCl2, 0.5 μL dNTPs, 0.5 μL Taq polymerase, 0.5 μL of each primer (aflR-1, aflR-2; omt-1, omt-2; ver-1, ver-2; and nor-1, nor-2), 0.5 μL template DNA, 1.5 μL deionized water; 14.5 μL was added to make the volume up to 25 μL. PCR technique was as follows: one step for denaturation at 96°C for 5 min followed by 35 cycles of: 95°C for 1 min; 65°C for 30 s; 72°C for 30 s; for the first cycle; and 94°C, 30 s; variable (56–62°C), 30 s; 72°C, 30 s for the next 34 cycles (Rashid et al., 2008).
For OTA assay, PCR reactions were performed in 50 μL as final volume, containing 5.0 μL of template DNA, 5.0 μL of 10×PCR buffer, 0.2 μM of each dNTP, 1.5 μM MgCl2, 0.3 μM each primer (ANPKS 1, 5′-CGACGACTAATCGCAAGTCA-3′ and ANPKS 2, 5′-CATCGTTGCTCATAGGGGTT-3′), and 2.5 U of Taq polymerase. The amplification process consisted of a pre-denaturation step at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C/30 s, annealing at 51°C/1 min and extension at 72°C/1 min, and a final extension of 7 min at 72°C (Castellá and Cabañes, 2011). Twelve microliters of the PCR products with positive and negative controls were loaded in the gel wells after mixing with appropriate amount of 6.7 μL of 6×loading dye. Also, 7.5 μL of 100-bp ladder mixed in 6×loading orange dye was loaded. PCR products were electrophoresed in a 1.2% agarose gel prepared in 1×TBE stained with ethidium bromide (1.5 μg/mL) for 2–3 h. DNA bands were visualized on a UV transilluminator at a wave length of 540 nm (Rashid et al., 2008).
Bioassay of toxins
The toxicity of A. flavus and A. niger strains were compared by bioassay against Artemia salina (Biji et al., 1981). The chloroform extract (0.05 mL) of fungal strains was placed in each test tube, the chloroform was evaporated, and about 10–20 shrimp larvae in 1 mL of salt water were transferred into tubes. The tubes were kept at room temperature (22–24°C). Control tubes with 0.05 mL of chloroform were also made. After 24 h, mortality was determined.
Results and Discussion
Date mycobiota
Data in Table 2 showed that 22 species belonging to 12 genera were collected from date fruits on 1% glucose–Czapek's agar at 28°C. The gross total counts of glucophilic fungi in dates samples were 1108 colonies per gram dry weight (Table 2). Most of the recovered fungi had been isolated previously from date fruits in many countries of the world (Nassar, 1986; Salik et al., 1999; Ragab et al., 2001; Gherbawy, 2001; Shenasi et al., 2002; Aba Alkhail et al., 2004; Alghalibi and Shater, 2004; Al-Sheikh, 2009; Bokhary, 2010), but almost all with different frequencies.
Calculated per gram dry weight of fruits.
Aspergillus, represented by five species, was the most frequently isolated genus. It was found in 45% of the samples comprising 30.4% of total fungi. In Egypt, Abdel-Sater and Saber (1999) found that Aspergillus was the predominant genus isolated from dates (100% of the samples). A. niger and A. flavus were the most prevalent species among aspergilla. They occurred in 36% and 72% of the samples, matching 4% and 20.8% of all cultured clones (Table 2). Abu-Zinada and Ali (1982) found that A. flavus was generally associated with some date fruit varieties from Saudi Arabia when the relative humidity (RH) was increased to 90% at 30°C. Nassar (1986) isolated Aspergillus represented by three species, A. niger and two species from Aspergillus (namely A. ruber and A. amstelodami) from dates in Aswan, Egypt. Shenasi et al. (2002) recorded A. flavus in 10 date fruit varieties from the United Arab Emirates (UAE) at the first stage of maturation. Alghalibi and Shater (2004) reported that Aspergillus was the most frequently isolated genus from date samples collected from Yemen and occurred in 75% of the samples comprising 71.4% of total fungi in dates. Also, they reported that Aspergillus niger was the most common Aspergillus species and isolated from 13 date samples out of 20.
Rhizopus stolonifer was the second most common fungus isolated in the present study. It occurred in 64% of the samples comprising 15.8% of total fungi (Table 2). Alghalibi and Shater (2004) isolated Rhizopus stolonifer from date palm fruits in Yemen, and it occurred in 25% of the samples comprising 11.7% of total fungi.
Penicillium was isolated from 27 samples of dates (54%). Similar findings were reported by Gherbawy (2001) in Egypt, who found Penicillium species isolated in high frequency from date fruit (53.3%). In contrast to those findings, Abdel-Sater and Saber (1999) found Penicillium species isolated in low frequency from dates (30%).
The remaining genera and species were isolated with low frequency and their total counts as represented in Table 2. Most of these fungi were isolated previously from date fruits in many parts of the world as reported by several researchers (Nassar, 1986; Salik et al., 1999; Ragab et al., 2001; Shenasi et al., 2002; Aba Alkhail et al., 2004; Al-Sheikh, 2009; Bokhary, 2010).
Aflatoxins and OTA assessment
Eighty isolates of A. flavus were subjected for detecting their aflatoxigenic potentials. Analysis of the toxigenic potentials of the tested isolates revealed that they varied in their abilities to produce total aflatoxins. Thus, although some isolates were toxigenic, others exhibited no detectable toxin production (Table 3). Seven out of 18 isolates showed aflatoxigenic potential, represented 38.88% of isolates collected. All the seven isolates showed aflatoxin B1 production; six isolates out of seven positive isolates showed aflatoxin B2 production, and only one isolate produced G1 and G2 (Table 3). The difference in production of diverse types of aflatoxins might be due to an inhibition by one (or all) of the parameters which alternatively were favorable for the production of aflatoxins. Contrary to those results, Mahjoub et al. (1989) reported that molds isolated from fresh and cold-stored Tunisian date variety Deglet Nour did not produce aflatoxins B1, B2, G1, and G2, sterigmatocystin, ochratoxin, penicillic acid, patulin, or citrinin. Toxigenic A. flavus and occasionally A. parasiticus were isolated from the surface of 12 Emirati date fruits at three different stages of maturation (Ahmed and Robinson, 1997). Abdel-Sater and Saber (1999) reported that aflatoxin B1 was found in two samples out of 20 tested samples of dry date in Egypt. Using TLC analysis of 40 different date samples, Ragab et al. (2001) revealed that two out of five samples of pitted date fruits stuffed with peanut were contaminated by aflatoxin B1. Alghalibi and Shater (2004) reported that two samples of dates in Yemen out of 20 samples were naturally contaminated with aflatoxin B1. All varieties of date fruits in Saudi Arabia tested by Aba Alkhail et al. (2004) were able to support the production of different forms of aflatoxins (B1, B2, G1, and G2), and the content of Aflatoxin G1 produced by either A. flavus or A. parasiticus were higher than the other forms.
T and NT, toxic and nontoxic for more than 50% mortality of larvae tested.
Thirty-six isolates of Aspergillus niger were tested for ochratoxin production (Table 4). The tested isolates showed variable abilities to produce ochratoxin. Only nine isolates (25%) were reported as ochratoxin producers (Table 4). Few previous investigations were interested in detection of ochratoxin producers in date fruits. In this respect, Abdel-Sater and Saber (1999) recorded a detectable amount of OTA in two samples of dates in Assiut, Egypt (360–450 μg/kg). Ferracin et al. (2009) tested 20 isolates from A. niger isolated from date fruits in Brazil for their ochratoxin potential. Their results indicated that only nine out of 20 isolates were ochratoxigenic.
T and NT, toxic and nontoxic for more than 50% mortality of larvae tested.
ND, not detectable by the TLC method; +, detectable by the TLC method.
Detection of the polymerase chain reaction (PCR) product on 1% agarose gel: −, no product detected; +, PCR product detected.
PCR assay
The results indicated that all aflatoxingenic strains of A. flavus (7 isolates) contained the four tested genes: norA, ver1, omtA, and avfA (aflR) genes (Fig. 1 and Table 3). These results complement the results of toxicity test and aflatoxin detection using TLC. Such results were in agreement with the findings obtained by Rashid et al. (2008), who stated that aflatoxigenic isolates from A. flavus and A. parasiticus had all four norA, ver1, omtA, and avfA (aflR) genes.
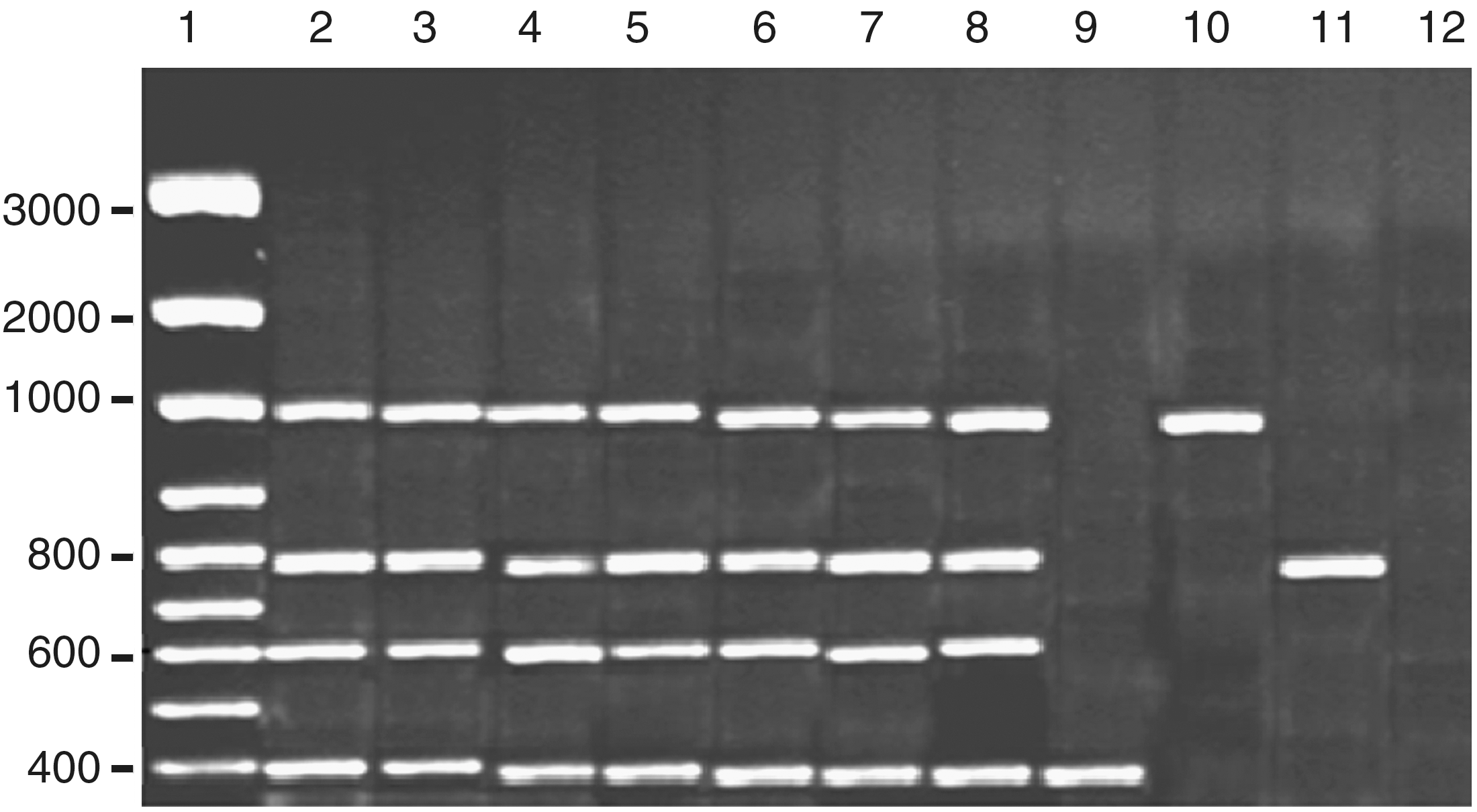
Amplification products obtained by quadruplex polymerase chain reaction (PCR) with the extracted DNA from the Aspergillus flavus isolates. Lane 1, DNA marker. Lanes 2–8 (TUAflavus1, TUAflavus4, TUAflavus5, TUAflavus10, TUAflavus11, TUAflavus12, and TUAflavus14) are toxigenic isolates. Lanes 9–11 (TUAflavus2, TUAflavus8, and TUAflavus17) are atoxigenic isolates. Lane 12, negative control.
Some of the atoxigenic isolates showed the presence of only one gene. For example, the three atoxigenic isolates TUAflavus2, TUAflavus8, and TUAflavus17 had norA, avfA, and omtA, respectively (Table 3). The aflatoxinogenic strains had a quadruplet pattern, indicating the presence of the four genes in the aflatoxin biosynthetic pathway, whereas nonaflatoxinogenic strains presented different patterns (Criseo et al., 2001). On the other hand, Yang et al. (2004) reported different results by multiplex-PCR and enzyme-linked immunosorbent assay (ELISA). They stated that, although norA, ver1, omtA, and avfA (aflR) genes had been detected in all tested strains, some of these samples were negative for aflatoxin detection. Gashgar et al. (2010) reported the presence of aflR1 gene in seven isolates of A. flavus isolated from wheat flour in Saudi Arabia.
Using the primer pairs of Castellá and Cabañes (2011), the ochratoxigenic isolates of A. niger isolated from date fruit samples were easily differentiated from nonochratoxigenic isolates (Table 4). The ochratoxigenic isolates produced a 951-bp DNA fragment with those primers, while no amplification was reported with nonochratoxigenic isolates (Fig. 2). Castellá and Cabañes (2011), using various PCR reactions, demonstrated that the identified fragment was very specific for OTA-producing strains of the A. niger aggregate. Also, the nonochratoxigenic strains of the A. niger aggregate gave negative results. Moreover, no PCR products were obtained with other ochratoxigenic species of Aspergillus section Nigri, such as A. carbonarius or A. sclerotioniger. All other fungi tested, even ochratoxin A–producing species from Aspergillus and Penicillium, proved to be negative.
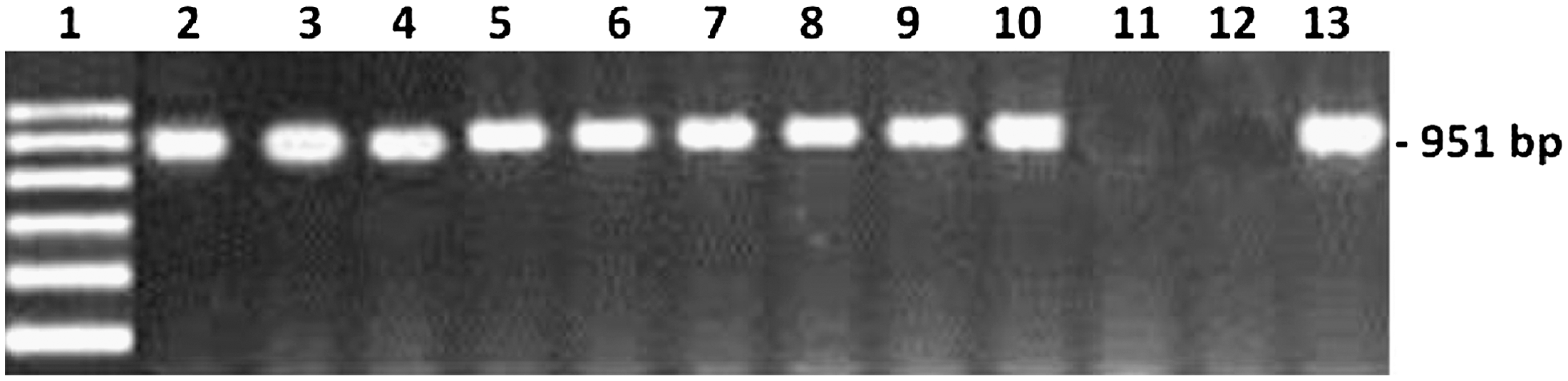
Specific detection of ochratoxigenic Aspergillus niger isolates using the primer pair ANPKS1and ANPKS2. Lane 1, DNA marker. Lanes 2–10, ochratoxigenic isolates (TUAniger2, TUAniger4, TUAniger5, TUAniger6, TUAniger10, TUAniger11, TUAniger12, TUAniger14, and TUAniger27). Lane 11, atoxigenic isolates (TUAniger1). Lane 12, negative control. Lane 13, positive control (KAUAnig2).
Footnotes
Acknowledgments
This work was supported by a grant (Contract No. 1024-432-1) from Taif University, Kingdom of Saudi Arabia.
Disclosure Statement
No competing financial interests exist.