
Abstract
The purpose of the present study was to evaluate and assess the capability of Zataria multiflora, Geranium herbarium, and Eucalyptus camaldolensis essential oils in treating Saprolegnia parasitica–infected rainbow (Oncorhynchus mykiss) trout eggs. A total of 150 infected eggs were collected and plated on glucose-pepton agar at 24°C for 2 weeks. The antifungal assay of essential oils against S. parasitica was determined by a macrodilution broth technique. The eggs were treated with essential oils at concentrations of 1, 5, 10, 25, 50, and 100 ppm daily with three repetitions until the eyed eggs stage. Of 150 eggs examined, S. parasitica (54.3%), Saprolegnia spp. (45%), and Fusarium solani (0.7%) were isolated. The minimum inhibitory concentrations of Z. multiflora, E. camaldolensis, and G. herbarium essential oils against S. parasitica were 0.9, 2.3, and 4.8 ppm, respectively. Zataria multiflora and E. camaldolensis at concentrations of 25, 50, and 100 ppm, and G. herbarium at concentration of 100 ppm had significant differences in comparison with negative control (p<0.05). The results revealed that malachite green, followed by Z. multiflora, E. camaldolensis, and G. herbarium treated eggs had remained the most number of final eyed eggs after treatment. The highest final larvae rates belonged to malachite green, E. camaldolensis, Z. multiflora, and G. herbarium, respectively. The most hatching rates were recorded with malachite green (22%), and then Z. multiflora (11%), E. camaldolensis (7%), G. herbarium (3%), and negative control (1%). Zataria multiflora and E. camaldolensis were more effective than G. herbarium for the treatment of S. parasitica-infected rainbow trout eggs in aquaculture environment.
Introduction
F
Malachite green is commonly used to solve this problem and has been widely reported as an effective fungicidal agent (Li et al., 1996). However, the use of malachite green on fish is banned in some countries because of its teratogenic effects and the finding of a residue called leucomalachite with carcinogenic activity, which can persist in fish tissues for a long time (Culp et al., 1999). Hydrogen peroxide, formalin, and sodium chloride at high concentrations are the other promising chemicals for controlling and the treatment of Saprolegnia infection with minimal impact to the environment (Fitzpatrick et al., 1995; Schreier et al., 1996; Celada et al., 2004; Racha et al., 2004; Melendre et al., 2006). Therefore, there has been significant effort expended to identify natural therapeutic agents, being effective as malachite green in combating mycotic infections of fish and fish eggs, but is safer for the operator, the fish, and the environment. Since the herbal essential oils were used in traditional folk remedies (Tadjbakhsh, 2003), previous studies have demonstrated their inhibitory effects on some pathogenic fungi (Khosravi et al., 2008). Several investigators have evaluated the possibility of using these natural products as fungicidal agents on fish eggs in aquaculture system (Mori et al., 2002; Rai et al., 2002; Tampieri et al., 2003; Chukanhom et al., 2005). The plants of Z. multiflora, G. herbarium, and E. camaldolensis, naturally occurring in central and southern Iran, indicated antifungal activity against different pathogenic fungi such as Candida, Aspergillus, and Fusarium species (Naeini et al., 2009; Shokri et al., 2011). There is no information concerning the efficacy of above-mentioned essential oils against aquatic fungi, in particular Saprolegnia species, in Iran. The goal of this project was to investigate three different herbal essential oils, including Z. multiflora, G. herbarium, and E. camaldolensis, as alternative options against saprolegniasis on rainbow trout (Oncorhynchus mykiss) eggs.
Methods
Water quality
Water flow was maintained at 12 L/min for each test tank. The mean water temperature was maintained within 11±2°C; dissolved oxygen concentration and pH were calculated approximately 8.1 ppm and 7.7 during treatment, respectively.
Sampling and isolation of infected eggs
A total of 150 fungus-infected rainbow trout eggs were obtained from several rainbow trout propagation farms in Yasoudje, Iran. Each sample, 5 eggs, was added to the universal tube (50 mL) containing sterile distilled water, hemp seeds, gentamicine (5 μg/mL), and chloramphenicol (16 μg/mL). They were transferred to Mycology Research Center in University of Tehran during 2 days by flasks with ice pack. In this study, all general chemical materials were purchased from Merck Co. (Darmstadt, Germany).
Isolation and identification of Saprolegnia and Fusarium
The eggs were plated on glucose-pepton agar and incubated at 24°C for 2 weeks. Saprolegnia isolates were identified based on morphological and growth features as previously described by Willoughby et al. (1985). The primary and secondary zoospores and cysts were observed as well as oogonia and antheridia. The secondary cyst had groups of long hairs (7–10 pm length), which were observed by light microscopy. They were germinated readily, forming septate germ tubes with the cytoplasmic material located in the distal area. A wild-type strain of S. parasitica obtained from the infected egg was used in this study.
Another part of infected eggs was cultured on potato dextrose agar (PDA) and incubated at 25°C for 5–7 days. All Fusarium isolates were subcultured on PDA, spezieller nahrstoffarmer agar (SNA), and carnation leaf agar (CLA). PDA cultures were incubated at 25°C for 5–7 days, and CLA and SNA cultures were incubated at 25°C for 2–4 weeks. Cultural characters were assessed by eye and by microscopic examination. Colony morphology was recorded from cultures grown on PDA. The morphology of macroconidia, microconidia, conidiogenous cells, and the chlamydospores was assessed from cultures grown on SNA and CLA. Morphological identifications of isolates were made using the criteria of Leslie and Summerell (2006).
Determination of minimum inhibitory concentrations (MICs) of essential oils
The plants of Z. multiflora (voucher no. 1106), G. herbarium (voucher no. 1319), and E. camaldolensis (voucher no. 1478) were chosen in this study. The samples (40 g of fresh material) were submitted to hydrodistillation for 3 h, using a Clevenger-type apparatus, according to the European Pharmacopoeia (1975). The volatile distillate was collected over anhydrous sodium sulphate and stored at −25°C in sealed glass vials until use. The in vitro susceptibilities of isolated S. parasitica against essential oils were determined by a macrodilution broth technique as previously described with modifications (CLSI, 2002). In this study, activity against the zoosporic stage was estimated with zoospores prepared in sterilized tap water at 10°C as described by Kitancharoen et al. (1996). Zoospores were harvested by aseptic filtration through gauze and adjusted to approximately 1×106 spores/mL using a haemocytometer slide. Dilutions of essential oils (diluted in 0.01% dimethyl sulfoxide) were serially prepared from stock solution in sabouraud dextrose broth ranging from 0.1 to 20 ppm. For the macrodilution broth test, 900 μL of the final spore suspensions were mixed with 100 μL of the test essential oils in 12×75 mm test tubes and incubated at 24°C for 48 h. The growth in each tube was compared with that of the growth control (essence-free) tube. The MIC was defined as the lowest concentration of essential oil, which produced no visible growth.
Exposure studies
The next step was to treat fungus-infected eggs with essential oils at different concentrations and exposure durations. In this regard, the eggs were randomly transferred into outdoor circular glass fiber stock tanks (40×40×15 cm), which were supplied with a constant flow of river water (12 L/min). Each tank contained approximately 4000 rainbow trout eggs. The parameters of water quality such as pH, dissolved oxygen, and temperature were recorded before and after the treatment.
The eggs were treated with different concentrations of Z. multiflora, G. herbarium, and E. camaldolensis essential oils (1, 5, 10, 25, 50, and 100 ppm). The essential oils were dissolved by pressure of river water, and no dispersant was added. During the incubation, the essential oils were administered in tanks with density of 4000 eggs daily with three repetitions for 1 h until the eyed eggs stage (about 18–20 days in 11°C water temperature). The oils were eliminated from the tanks by using hot water plus Savlon (Novartis, Horsham, UK) in different times. The treatments were compared to negative control, which received no treatment, and to that treated with malachite green, which was used as positive control. The malachite green group was treated with 1 ppm of this agent, and its treatment was performed the same as in other groups. All the treatments were carried out in triplicate. In all experiments, the fungal degree, the hatching rate, the final larvae rate, and the mean larvae weight were calculated at days 1, 5, 10, 15, and 18. The fungal degrees were expressed as fungal colony counts. Egg hatching rates were calculated from the incubated buoyant eggs and expressed as percent of the buoyant eggs. In addition, final larvae rates were measured as the number of larvae obtained from the initial number of eggs, and the mean larvae weights were calculated by weighing the exact number of larvae resulted in all larvae weights. The data were analyzed by one-way analysis of variance, followed by post-hoc test (Bonferroni) and Pearson correlation test using SPSS (version 11). A p-value of less than 0.05 was statistically considered significant.
Results
Presence of Saprolegnia and Fusarium
Of 150 eggs examined, S. parasitica (54.3%) was the most prevalent fungi, followed by Saprolegnia spp. (45%), while Fusarium solani was isolated only from one sample (0.7%).
MIC values of essential oils
Among the isolated fungi, S. parasitica (SIR-22) was randomly chosen for evaluating the antifungal effect of essential oils. The in vitro susceptibility test indicated that S. parasitica was more susceptible to Z. multiflora (0.9 ppm) than E. camaldolensis (2.3 ppm) and G. herbarium (4.8 ppm).
Comparison of essential oils and malachite green as antifungal agents
All treatments demonstrated increasing fungistatic properties with increasing concentrations, but there were no remarkably effect in some doses, for example, Z. multiflora at concentrations of 25 and 50 ppm. As shown in Figure 1, there were statistically significant differences among concentrations applied in treatment groups in inhibiting the fungal growth (p<0.05). Positive control had the lowest mean fungal degrees (6.5), followed by E. camaldolensis (42) and Z. multiflora (43.5), while negative control and G. herbarium had the highest mean fungal degrees in values of 113.5 and 71.39, respectively. When comparing hatching rates, there were highly significant differences between the treatments and negative control (p<0.05). Positive control and Z. multiflora had the highest mean hatching rates in values of 22% and 11%, respectively, while negative control had the lowest hatching rate with a mean value of 1%. The hatching rates were 7% for E. camaldolensis and 3% for G. herbarium (Table 1).
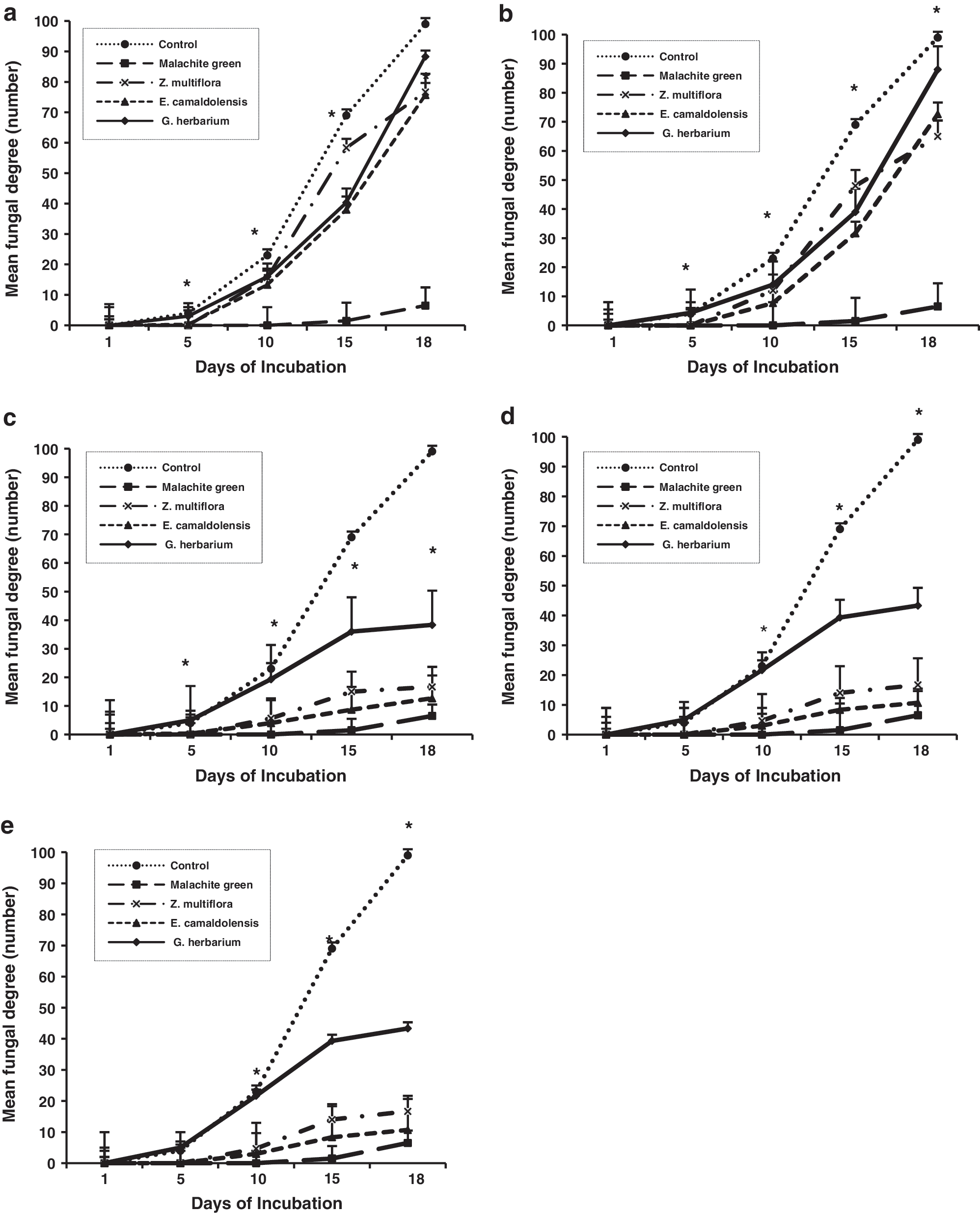
Mean fungal degrees of treatments at 5 ppm
p<0.05 (significant differences).
Data analysis showed that Z. multiflora and E. camaldolensis had the highest eyed egg rates and final larvae rates with mean numbers of 1651.61 and 310.66, respectively. Significant differences were observed among the essential oils and controls (p<0.05), while there were no statistically significant differences among the essential oils tested. Regarding the mean larvae weight rates, statistically significant differences among the treatments were not detected (Table 1).
Variance analysis showed significant differences based on fungal degrees between Z. multiflora and E. camaldolensis at concentrations of 1 ppm at day 15, 5 ppm at day 5, 10 ppm at day 15, 25 ppm at day 15, and 50 ppm at days 15 and 18, between E. camaldolensis and G. herbarium at concentrations of 10 ppm at day 5, 50 ppm at day 5, and 100 ppm at day 5, between Z. multiflora and G. herbarium at concentrations of 5 ppm at days 5 and 18, 10 ppm at day 5, 25 ppm at day 5, 50 ppm at day 5, and 100 ppm at day 5 (p<0.05). Totally, the effects of the essential oils tested on different stages of incubated eggs were as follows: in lowering fungal degree, malachite green > Z. multiflora > E. camaldolensis > G. herbarium > negative control; in hatching rate, malachite green > Z. multiflora > E. camaldolensis > G. herbarium > negative control; in eyed eggs stage, malachite green > Z. multiflora > E. camaldolensis > G. herbarium > negative control; in final larvae rate, malachite green > E. camaldolensis > Z. multiflora > G. herbarium > negative control; in yielding total larvae weight, malachite green > E. camaldolensis > Z. multiflora > G. herbarium > negative control. Among the different essential oils used, Z. multiflora had more effect in controlling the fungal growth, hatching percent, and the eyed eggs than the other ones.
Discussion
This study was performed in circular glass fiber stock tanks, which were normally used to hatch and to treat eggs. The essential oils extracted from three plants, including Z. multiflora, E. camaldolensis, and G. herbarium, were tested to know whether they inhibit the growth of S. parasitica (at concentrations of 1, 5, 10, 25, 50, and 100 ppm). To our knowledge, this study was the first project on these herbal essential oils in inhibiting the growth of S. parasitica.
In our study, all the essential oils demonstrated fungistatic properties against S. parasitica. By comparing in vitro and in vivo results, we obtained good results with Z. multiflora in vitro and E. camaldolensis in vivo in inhibiting the growth of S. parasitica. Although the in vitro and in vivo results were different, these differences were not statistically significant. This finding is in agreement with other drugs and chemical agents tested by some investigators (Yuasa and Hatai, 1995, 1996). The main components of Z. multiflora (thymol and carvacrol), E. camaldolensis (anethole and 1,8-cineole), and G. herbarium (geraniol and citronellol) have been associated with antimicrobial activities (Kalemba and Kunicka, 2003), and the possible mechanisms of action of them appear to be degradation of the cell wall, damage to cytoplasmic membrane, damage to membrane proteins, leakage of cell contents, and depletion of the protein motive force (Burt, 2004).
Data analysis showed that Z. multiflora at concentration of 25 ppm, E. camaldolensis at concentration of 25 ppm, and G. herbarium at concentration of 100 ppm for 60 min daily with three repetitions were the best treatments, representing the prevention of fungal attack, the increase of hatching rate, the eyed egg rate, and final larvae rate. Zataria multiflora and E. camaldolensis treatments showed positive significant differences in comparison with G. herbarium and negative control group (p<0.05), while G. herbarium treatment indicated negative significant differences in comparison with Z. multiflora, E. camaldolensis, and malachite green. In controls, malachite green (positive control group) had positive significant differences in comparison with G. herbarium and negative control (p<0.05), and negative control showed positive significant differences in comparison with Z. multiflora, E. camaldolensis, and malachite green (p<0.05). Although literature search showed that there is no research about antifungal activity of these tested oils against S. parasitica, some investigators demonstrated the inhibitory effects of different plants on some fungi in the saprolegniaceae such as Artemisia verlotorum, and Santolina etrusca extracts against S. ferax (Macchioni et al., 1999), Blumea balsamifera, B. mollis, Eupatorium triplinerve, Guizotia abyssynica, and Tagetes erecta essential oils against S. ferax (Rai et al., 2002) and Alpinia galanga essential oil against S. parasitica, S. diclina, Achlya bisexualis, A. diffusa, and Aphanomyces piscicida (Chukanhom et al., 2005). Inhibitory effects of essential oils of aegle, ageratum, citronella, eucalyptus, geranium, lemongrass, orange, palmarosa, patchouli, peppermint, and clove against fungal strains isolated from fish eggs were reported by other investigators (Pattnaik et al., 1996; Keene et al., 1998) as well.
Several studies showed that malachite green has proved to be an excellent antifungal agent in fish eggs at concentrations of 1–5 ppm (Kitancharoen et al., 1996, 1998), which were in close agreement with the results of our experiment. All the essential oils had lower activity than that of malachite green. In this regard, the dispersion of hydrophobic components of essential oils in the tank water is the main problem in oils activity testing, which lowers their activity. Additionally, a variety of other factors, such as of solvent or dispersing agent and aerobic and anaerobic growth conditions, can influence the activity of essential oils (Kalemba and Kunicka, 2003). Among the essential oils tested, Z. multiflora had higher activity than the other ones in most of stages. Based on previous studies, essential oils with phenols, for example, Z. multiflora, as main compounds express the highest activity against microorganisms, and their activity spectrum is the broadest, while oils with ethers (E. camaldolensis) and alcoholic (G. herbarium) compounds are slightly less active (Tampieri et al., 2003). By using a concentration of 1 ppm of Z. multiflora and E. camaldolensis essential oils applied in our study, the results didn't statistically differ from the 1 ppm malachite green at the same frequency. Because of higher activity of the two above essential oils at concentration of 25 ppm than that of 1 ppm and the banning of green malachite, we recommend that, in addition to routine screening of dead eggs, treatments with Z. multiflora and E. camaldolensis essential oils are preferred and they are good alternatives to improve the fungal infection and increase the hatching rate of rainbow trout eggs. In our study, the hatching rates are low and it may be due to the toxicity of essential oils tested, so further research must be carried out in order to determine the toxicity of essential oils as well as to test new natural fungicides in the future.
Footnotes
Acknowledgments
This work was supported by the Research Council of the University of Tehran, Iran.
Disclosure Statement
No competing financial interests exist.