
Abstract
Four nucleic acid extraction methods were evaluated for the purpose of quantifying a norovirus surrogate (murine norovirus [MNV-1]) concentrated from different food samples. Simple (strawberries and lettuce) and complex (sliced turkey breast, soft-shell clams, and potato salad) food matrices were inoculated with a viral suspension containing high (4×105 PFU) or low (4×103 PFU) numbers of viral particles. MNV-1 was eluted using either the Pulsifier™ or repetitive pipetting. The four methods were based on using magnetic silica (MiniMAG), non-magnetic silica (bioMérieux Basic kit), silica membrane (Qiagen kit), and phenol (TriReagent) for RNA extraction. The greatest recovery of viral RNA from simple matrices was obtained using magnetic silica for both inoculation levels. For strawberries, the addition of pectinase during the elution step improved RNA recovery when the Pulsifier was used with silica membrane extraction and when repetitive pipetting was used with magnetic silica extraction. In the case of complex matrices, the extraction of high or low numbers of MNV-1 was highest overall using magnetic silica. The exception was soft-shell clams with a high viral load, in which the greatest recovery was obtained with the phenol-based method. In general, magnetic silica was the most effective for extracting both high and low numbers of MNV-1 particles from a wide range of foods.
Introduction
Ready-to-eat (RTE) foods are now recognized as a major vehicle for norovirus (NoV) transmission, of which there have been international outbreaks involving multi-ingredients foods (40.2%), produce (16.5%), seafood (13.0%), and bakery items (8.7%), among others (Greig and Ravel, 2009). NoVs are responsible for over 90% of all viral gastroenteritis and about 50% of all outbreaks of foodborne illness worldwide (Patel et al., 2009). Since human NoVs are non-cultivable, surrogates such as feline calicivirus and murine norovirus are widely used to examine the behavior, persistence, and inactivation of human NoVs (Cannon et al., 2006). Due to their presence in low numbers, NoV detection in foods is problematic, which makes the monitoring and containment of outbreaks involving NoVs difficult. Different studies have shown that molecular methods can be used to detect NoVs in foods (D'Souza and Jaykus, 2006; Jean et al., 2004; Pinto and Bosch, 2008). However, the complexity of food matrices and the presence of enzyme inhibitors, which are co-extracted during viral concentration steps, cause considerable variation in the threshold of detection (D'Souza and Jaykus, 2006). Furthermore, the efficiency of extraction methods as well as the presence of potential inhibitors affect the reverse transcriptase–polymerase chain reaction (RT-PCR) reaction (Martin-Latil et al., 2012). Process or internal controls must be used to monitor the efficiency and to identify false-negative results especially for unknown viral load. Thus, there is a need for viral extraction methods that efficiently remove agents which interfere with the molecular detection of NoVs, suitable for different types of foods and are inexpensive. The objective of the present study was to compare four different RNA extraction methods in the context of NoV detection in RTE food by real-time RT-PCR.
Materials and Methods
Viral stock
Murine norovirus (MNV-1) was propagated in RAW 264.7 cells as previously described (Wobus et al., 2004). The cell line and virus were kindly provided by Kirsten Mattison (Health Canada, Ottawa, ON). The titer of the MNV-1 stock suspension was estimated at 2×107 PFU/mL based on a plaque assay (Girard et al., 2010).
Food matrices
For the extraction of viral RNA, lettuce and strawberries were considered as simple food matrices, while turkey breast, potato salad, and soft shell clams were considered as complex food matrices. All products were purchased in a local supermarket and inoculated with 200 μL of viral suspension (diluted to 2×104 or 2×106 PFU/mL). For all the samples, MNV-1 was allowed to attach for 5 min under a laminar flow hood. A short attachment time was used based on previous studies (Girard et al., 2010; Rzezutka et al., 2005). Samples were prepared as follows:
Lettuce (romaine type)—Green portions (4×4 cm for a total weight of 10 g) were washed twice with 10 mL of distilled water, three times with de-ionized water, and then dried for 35 min under a laminar flow hood before adding MNV-1.
Strawberries (freshly harvested, Île d'Orléans, Québec, Canada)—A total weight of 10–13 g was washed as described for lettuce.
Turkey breast—10-g slices (4×4 cm) were inoculated with MNV-1 as described for lettuce.
Potato salad (containing potatoes, mayonnaise, and herbs)—10-g portions of potato cubes were inoculated with MNV-1 as described above.
Soft-shell clams—Clams were harvested in the Capucins region (Gaspésie) near Saint-Omer, Québec, Canada; the digestive glands were removed and inoculated with MNV-1 as described above.
Elution and concentration of the norovirus surrogate
Two methods were used to elute the viral particles from the different food matrices: Pulsifier™ (Microbiology International, Frederick, MD) and repetitive pipetting. The Pulsifier treatment was performed as described previously (Trottier et al., 2006). The repetitive pipetting treatment was carried out as described earlier (Simard et al., 2007). For both methods, the PEG pellet was resuspended in 180 μL of phosphate-buffered saline (PBS; for methods with silica) or 2.5 mL of TriReagent (for the phenol-based method).
In order to look for an improvement on the recovery of MNV-1 particles from strawberries, some samples were gently mixed with 100 μL of Pectinex Ultra SPL (Sigma-Aldrich Canada, Ltd., Oakville, ON, Canada). Samples were incubated at room temperature for 30 min prior to the PEG concentration step.
Nucleic acid extraction methods
All RNA was stored at −80°C or used immediately for molecular detection after extraction by one of the following methods:
Non-magnetic silica—Viral RNA was extracted using the Basic extraction kit (CAT 284123) according to the manufacturer's instructions (bioMérieux, Marcy l'Etoile, France). Briefly, the PBS suspensions were mixed with 0.9 mL of lysis buffer and 50 μL of silica. After centrifugation for 2 min at 13,000 rpm (MiniSpin plus; Eppendorf, Hamburg, Germany), the pellet was washed with 500 μL of wash buffer, then with 1 mL of 70% ethanol and finally with 1 mL of acetone. Samples were dried for 10 min at 56°C, then mixed with 50 μL of elution buffer and incubated 10 min at 56°C. RNA was finally recovered by centrifugation at 13,000 rpm for 2 min.
Phenol-based extraction—The phenol-based nucleic acid extraction was carried out according to the Health Canada Compendium of Analytical Methods (OPFLP-01 and OPFLP-03). Briefly, after concentration of the virus using PEG, the pellet was resuspended in 2.5 mL of TriReagent (Sigma-Aldrich) instead of 180 μL of PBS. After successive washes (chloroform, isopropanol, and ethanol), RNA was mixed with Dynabeads Oligo-(dT)25 (Invitrogen, Burlington, ON) in binding buffer. Beads were washed three times with washing buffer, and polyA-RNA was eluted using 50 μL of elution buffer at 90°C for 2 min.
Magnetic silica—Viral RNA was extracted using the NucliSens MiniMAG extraction kit (CAT 200293) according to the manufacturers' instructions (bioMérieux). Briefly, the concentrated viral particles in PBS were mixed with 1 mL of lysis buffer. Then 50 μL of silica was added and later centrifuged at 13,000 rpm (MiniSpin plus, Eppendorf) for 2 min. The pellet was washed twice with 400 μL of “wash buffer 1” and twice with 500 μL of “wash buffer 2.” The final wash was carried out with 500 μL of “wash buffer 3.” RNA was eluted with 50 μL of elution buffer at 65°C for 5 min.
Silica-based membrane—Viral RNA was extracted using the QIAamp MinElute Virus Spin kit (CAT 57704) according to the manufacturers' instructions (Qiagen Sciences, Gaithersburg, MD). Briefly, the eluted MNV-1 particles were mixed with 25 μL of protease (included in the kit) and 200 μL of buffer AL (containing 28 μg/mL of carrier RNA). The mixture was held at 56°C for 15 min. Absolute ethanol (250 μL) was added, and the column was held at room temperature for 5 min and then centrifuged at 8,000 rpm for 1 min. The column was washed successively with 500 μL of buffer AW1, 500 μL of buffer AW2, and 500 μL of ethanol. After drying, RNA was eluted from the column with 30 μL of buffer AVE.
Real-time RT-PCR (TaqMan)
MNV-1 RNA was amplified according to the method described by Girard et al. (2010) and using the MGmnv-1Forward primer (GCTGCGGCCTCTCTTGAC), MGmnv-1Reverse primer (AGGGATGGTGTCCTGAAAACC), MNV-1Probe (6FAM-TTCGTGCGGTCCCAAGATCCATCT-TAMRA), and AgPath ID RT-PCR kit (Ambion, Austin, TX). A standard curve was constructed using 10-fold serial dilutions of RNA extracted directly from the MNV-1 stock suspension using the magnetic silica method. Samples showing cycle thresholds (Ct) values of ≤36, with no evidence of amplification in the negative controls, were considered positive. Reaction efficiency was calculated using the formula E=(10−1/slope)-1 (Rasmussen, 2000). Quantification was performed in triplicate, except for the soft-shell clam samples which were measured in quadruplicate due to the complexity of the matrix. The viral recoveries (%) were calculated as follows: [(PFU/mL×0.2 mL×10) / (4×103 or 4×105)]×100.
Statistical analysis
Statistical analyses were performed with GraphPad Prism 5 (GraphPad Software Inc., La Jolla, CA). Significant differences (p<0.05) among mean values were tested by analysis of variance (ANOVA) and Bonferroni's Multiple Comparison test.
Results
Quantification of viral RNA
The reaction efficiency of the real-time RT-PCR assay used to calculate all recoveries was 0.95. The standard curve was determined as follows: y=−3.4399x+34, with R2=0.997.
Extraction of viral RNA from samples treated with the Pulsifier
No significant difference (p<0.05) was observed between the RNA extraction methods using non-magnetic or magnetic silica in the case of strawberries contaminated with a high viral load (Table 1). A difference of more than 2 log10(PFU/mL) was observed between the magnetic silica and phenol-based extraction methods for testing strawberries with a high viral load. No RNA was detected in strawberries with either inoculation level using the silica-based membrane. More RNA was recovered from lettuce, at both viral loads, using the magnetic silica-based method than the other methods (Table 1). Extraction using either magnetic or non-magnetic silica particles gave the best RNA recoveries with the lower viral load. Recovery was also higher in the case of sliced turkey breast with the higher viral load (Table 2). RNA recoveries from potato salad were not significantly different among both silica particle based methods and the phenol-based method. Unfortunately, the silica-based membrane was clogged by the potato salad sample, and thus values were not determined. There was no significant difference in RNA recovery between the phenol-based and magnetic silica extractions for the low viral load, although the amount of particles recovered using phenol extraction was greater (p=0.9388).
From a 200-μL suspension of MNV-1 applied to a food surface and dried. Values are averages of quadruplicate experiments. Values with different letters are significantly different for each food at p<0.05.
From a 200-μL suspension of MNV-1 applied to a food surface and dried. Values are averages of quadruplicate experiments. Values with different letters are significantly different for each food at p<0.05.
Viral recovery is expressed as a percentage of the initial load in Figure 1. The magnetic silica-based method gave recoveries of 6% and 3.7% of the high load in the cases of lettuce and strawberries, respectively. For the lower load, recoveries of 3.7% and 6.2% were obtained. Using the magnetic silica-based extraction from sliced turkey breast yielded higher recoveries of 46.6% and 51.2% for both low and high loads, respectively. In the case of potato salad and soft-shell clams, recoveries of <1% were obtained for both low and high loads using magnetic silica.
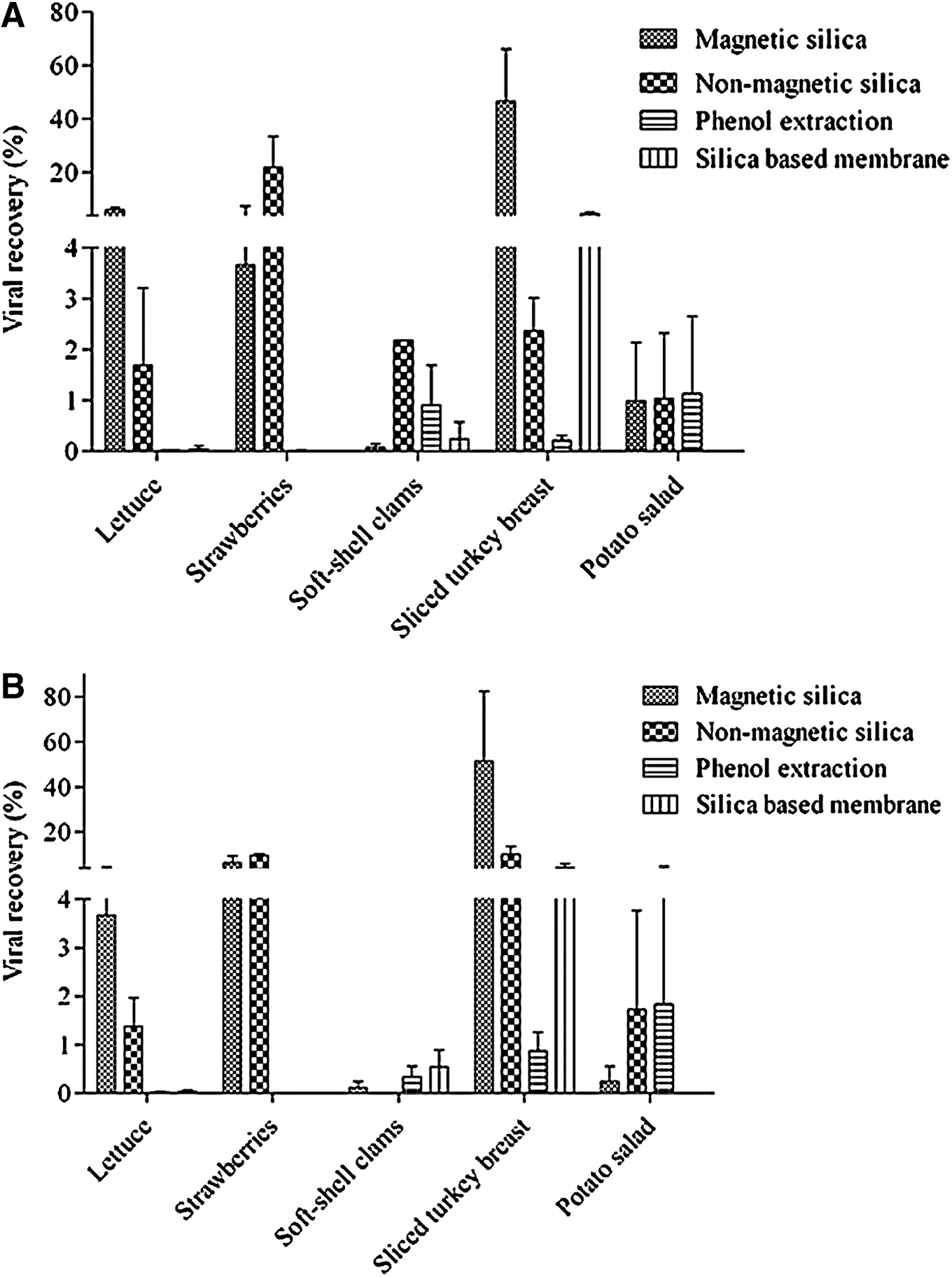
Percent recovery of high
Extraction of viral RNA from samples treated using the pipette technique
Only results obtained using the silica-based membrane and magnetic silica particles are shown (Table 3), since the non-magnetic silica was discontinued by the manufacturer and was no longer available at the time of these experiments. Furthermore, the phenol-based extraction method worked poorly in conjunction with the Pulsifier. Magnetic silica was more effective on sliced turkey breast and lettuce for the higher viral load, while the silica membrane gave the best result in the case of strawberries.
From a 200-μL suspension of MNV-1 applied to a food surface and dried. Values are averages of triplicate experiments Values with different letters are significantly different for each food at p<0.05.
Effect of pectinase during viral elution on subsequent extraction of RNA
In the absence of pectinase, processing strawberries with the Pulsifier led to clogging of the Qiagen column membrane, thus interfering with viral RNA extraction. The addition of pectinase (100 μL, 30 min at room temperature) during this step enabled the subsequent extraction of RNA, for both the high and low viral loads, with the Qiagen kit (Fig. 2). In comparison, the use of pectinase in conjunction with the pipette elution technique and the magnetic silica-based method strongly enhanced subsequent RNA extraction up to 6% and 3% for the high and low viral loads, respectively. Viral recovery was doubled with the use of pectinase in conjunction with the Pulsifier and magnetic silica-based methods (Fig. 2).
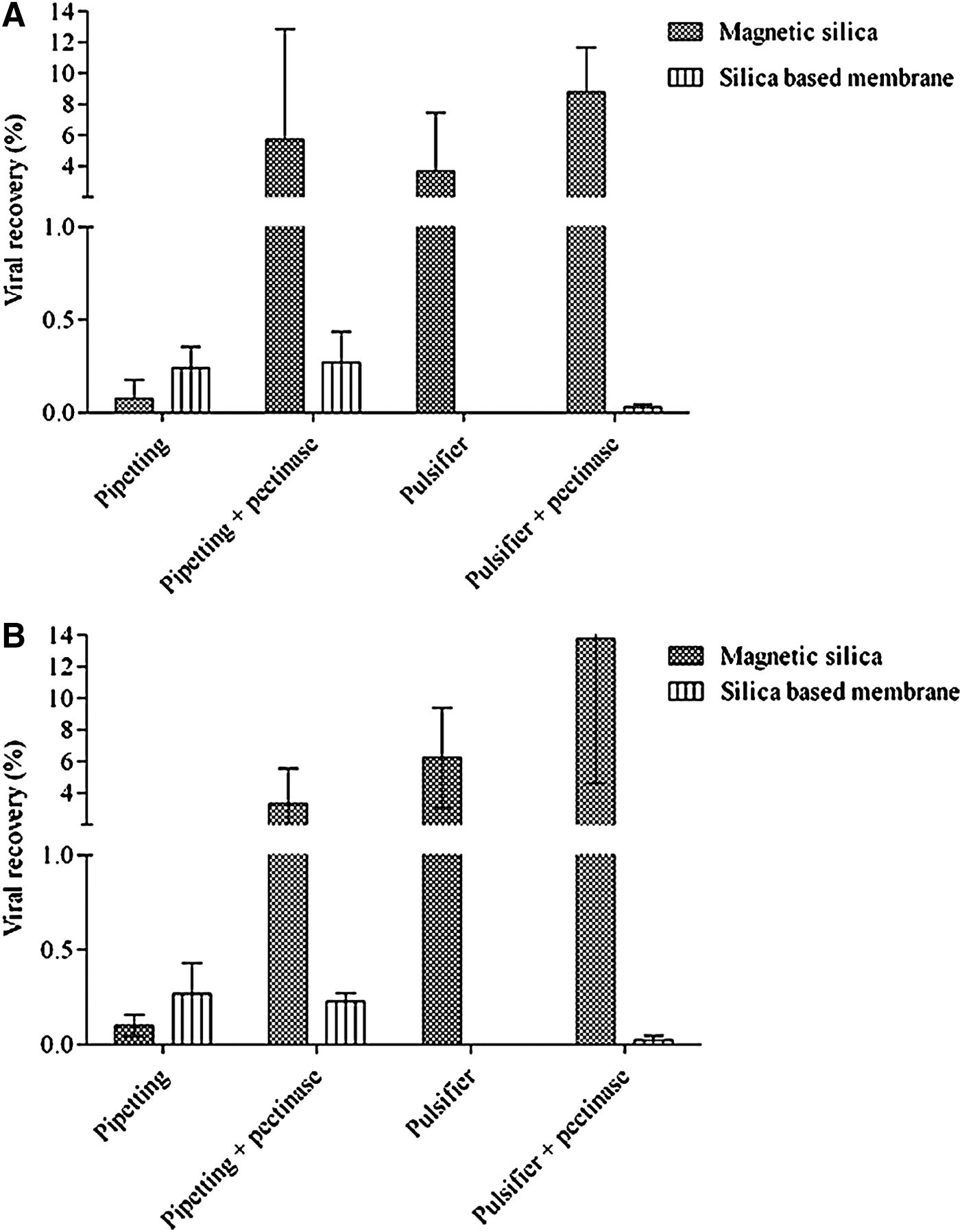
Elution methods for recovery of high
Discussion
The present study investigated the possibility that one RNA extraction method could be suitable for both simple and complex RTE food matrices. The high binding of viral RNA by magnetic silica from strawberry and lettuce, for both inoculation levels tested, showed the suitability of this method for simple matrices. Contrary to previous studies (Baert et al., 2008; Bianchi et al., 2011; Kim et al., 2008), the use of a silica-based membrane did not represent an improvement in the recovery of viral RNA from complex matrices in this study. This discrepancy might be due to differences in sample preparation methods, as strawberry homogenates (Bianchi et al., 2011) or 5-h shaking of whole strawberries (Kim et al., 2008) were used previously. In the present study, the use of Pulsifier for the elution of viral particles helped to minimize the disruption of the sample matrix. In a recent study, high viral recoveries were obtained using a Pulsifier in the case of fresh-cut vegetables (Sanchez et al., 2012). However, the release of some strawberry components, such as pectin, into the primary solution might have been higher and consequently, the silica membrane was clogged. Higher recoveries of NoV using a silica membrane were reported in the cases of grapes, strawberries and raspberries (Kim et al., 2008), although a long elution method was employed, which might not be applicable to more complex matrices such as potato salad or turkey breast. Complex food matrices generally contain more fat and protein than fruits and vegetables and are therefore more difficult to analyze (Baert et al., 2008). The present results show that silica particles are able to recover RNA from primary solutions of complex food matrices such as sliced turkey breast and potato salad. The use of non-magnetic rather than magnetic silica may improve the quantification of viral RNA from soft-shell clams, although only one of four samples provided a detectable response. Comparable results were obtained previously with oysters using a non-magnetic silica-based method (Gentry et al., 2009). The present findings suggest more non-specific binding and lower biocompatibility between the clam components and the magnetic silica particles. Furthermore, the more specific interaction between the viruses and the receptors present in clams might also have influenced the viral elution and, consequently, the RNA recovery. Other possible explanations for the difference in the recovery between magnetic and non-magnetic silica are the unbinding of RNA and the formation of aggregates. Compared to the non-magnetic particles, the magnetic silica particles may have a greater tendency to form complex aggregates with genomic DNA and/or non-viral RNA in the clam pellet. The formation of aggregates between large nucleic acid molecules and silica particles has been observed since the establishment of the Boom method (Boom et al., 1990). Finally, since organic molecules do not form strong chemical bonds (such as covalent bonds) with magnetic particles, the desorption and loss of viral RNA during the washing steps is possible (Kalele et al., 2006).
In addition to having the highest RNA recovery with a high viral load, the magnetic silica was the most efficient at extracting RNA from turkey breast and potato salad loaded with low viral counts. It was observed in previous studies that a silica based membrane is highly suitable for RNA extraction of low amounts of MNV-1, either from viral suspensions (Morales-Rayas et al., 2010) or from simple matrices such as strawberries and lettuce (Morales-Rayas et al., 2009). Nevertheless, a silica-based membrane was not suitable for RNA extraction from complex matrices such as mussels (Morales-Rayas et al., 2009). This is in accordance with our results, as detection of MNV-1 was not possible from potato salad, but low amounts were detected in sliced turkey breast and soft-shell clams. It was also found that silica-based membrane was not suitable for the detection of MNV-1 in three different pork matrices (Martinez-Martinez et al., 2011) The use of Pulsifier in the present study may have influenced the performance of the silica-based membrane, since a low amount of food debris was produced in the soft-shell clam primary solution. A phenol-based extraction combined with magnetic separation was shown to perform poorly overall in the present study, suggesting the unsuitability of this method for purification of viral RNA from simple RTE foods. Previous studies with hepatitis A virus in strawberry homogenates (Bianchi et al., 2011), NoV GII.4 in whole strawberries (Boxman et al., 2007; Kim et al., 2008), and on lettuce, fruit salad, and raspberries (Baert et al., 2008) support the poor performance of this type of RNA extraction from simple matrices. The presence of inhibitory substances such as phenolic compounds (Stals et al., 2011; Wilson, 1997) may have compromised the phenol-based method. This method was designed primarily to separate fat and protein, and therefore, it is commonly applied to complex matrices when high food particulates and components such as proteins, fat, and carbohydrates should be removed (Rutjes et al., 2006).
For both inoculation levels tested, Pulsifier and magnetic silica-based method resulted in viral RNA recoveries from strawberries of less than 10% of the initial viral load. Due to the higher RNA recoveries from simple matrices such as strawberries, another elution method (repetitive pipetting) was tested on the three matrices (turkey, lettuce, and strawberries) that yielded higher recovery using the Pulsifier. The similar recoveries from sliced turkey breast and lettuce when using magnetic silica and repetitive pipetting still supported the use of magnetic silica. However, repetitive pipetting elution combined with the silica membrane extraction improved the RNA viral recovery from strawberries, making MNV-1 detectable with the molecular assay. The differences in the viral recovery between the two elution methods might have been due to the Pulsifier effect on the soft structure of the strawberry. The high oscillation speed may destroy the soft structure of this fruit, which could explain the poor performance of the silica membrane using this elution method. The addition of pectinase in the primary solution from the fruit improved the subsequent RNA extraction by the magnetic silica as well as the silica membrane method when elution included use of the Pulsifier. Nevertheless, the highest recoveries were obtained by using the magnetic silica to purify the viral RNA.
Considering the present results, extraction based on magnetic silica (MiniMAG) seems most adequate for recovering RNA from complex and simple RTE food matrices with different viral loads.
Footnotes
Acknowledgments
We thank Andrée-Anne Martel for technical assistance. We also thank bioMérieux for providing the different kits used in this study. The financial support provided by the Natural Sciences and Engineering Research Council of Canada (NSERC) and Le Fond Québécois de la Recherche sur la Nature et les Technologies (FQRNT) is also acknowledged.
Disclosure Statement
No competing financial interests exist.