
Abstract
It is well documented that stress-adapted microorganisms can develop cross-resistance to other unrelated stress. This study was designed to evaluate the thermal resistance of acid-adapted Escherichia coli O157:H7 in both fresh and finished dairy composts. A three-strain mixture of E. coli O157:H7, either acid-adapted or non-adapted (control), was inoculated into dairy compost to a final concentration of approximately 107 CFU/g. The inoculated compost was kept in an environmental chamber which was programmed to raise temperature from room to target temperatures (50°C, 55°C, and 60°C) in 2 days, simulating the early phase of composting. In fresh dairy compost with 2 days of come-up time, acid-adapted and control E. coli O157:H7 survived for 19 and 17 days at 50°C, respectively, and 6 and 4 days for both types of culture at 55°C and 60°C, respectively. Overall, pathogen survival was non-significant (p>0.05) between control and acid-adapted cultures at all tested temperatures. In finished compost, the same trend in pathogen survival between control and acid-adapted cultures was observed at 55°C. However, the duration of survival for both cultures was short in comparison to that in fresh compost. In fresh compost with short come-up time (15 min), acid-adaptation provided E. coli O157:H7 some cross-protection to heat at 55°C up to 30 min of exposure. The effect of heating medium on thermal resistance of acid-adapted E. coli O157:H7 revealed that in saline, acid-adapted E. coli O157:H7 was inactivated slower (p<0.05) with 0.5 and 1 h of heat exposure at 55°C as compared to control culture. Our results revealed that cross-protection against heat in E. coli O157:H7 due to acid-adaptation was demonstrated in saline but lost in fresh dairy compost with 2 days of come-up time during composting. Additionally, the type of compost and heating medium can influence the rate of pathogen inactivation at composting temperatures.
Introduction
Besides heat-shock response, acid or low-nutrient (starvation) stresses are also known to provide protection to microorganisms through cross-protection mechanisms (Buchanan and Edelson, 1999; Rowe and Kirk, 2000). General stress proteins such as DnaK and GroEL are also induced during acid-adaptation and provide cross-protection to bacterial cellular components (Heyde and Portalier, 1990; Rodriguez-Romo and Yousef, 2005). These protection mechanisms may prolong pathogen survival in compost, thereby leading to contamination of crop and produce.
Acid-adaptation of E. coli O157:H7 can happen when the microorganism is exposed to gastric acidity and volatile fatty acids produced in the rumen and the intestine during the normal course of animal production (Giannella et al., 1972). The extent of acid-adaptation can increase during grain feeding of cattle due to low pH in rumen and production of several acids in the gastrointestinal tract, whereas during forage feeding this effect is mild (Diez-Gonzalez et al., 1998; Callaway et al., 2003). Acid-adaptation of E. coli O157:H7 can increase their survival time when exposed to heat or further acidic conditions (Ryu and Beuchat, 1998; Cheng et al., 2003). This adaptive response could be a problem in compost as it potentially extends pathogen survival beyond 3 days at 55°C (composting guidelines of USEPA). Under optimal composting conditions, E. coli O157:H7 is inactivated in 2 days (Singh et al., 2011); however, under suboptimal composting conditions, it has been shown to survive for up to 60 days (Shepherd et al., 2010). Therefore, the objective of this study was to investigate the effect of acid-adaptation on thermal inactivation of E. coli O157:H7 in dairy compost by simulating early phases of composting process under controlled environmental conditions.
Methods
Compost preparation
Fresh dairy compost with optimal carbon to nitrogen (C:N) ratio of 24:1 was prepared as previously (Singh et al., 2011). The fresh compost mix was analyzed by the Agricultural Service Laboratory (Clemson University, Clemson, SC) for the C:N ratio. The compost mix thus made was stored under refrigeration condition and used within 2 weeks.
At 1 day before the experiment, the moisture content of the compost was adjusted to approximately 50% and measured as previously (Singh et al., 2011).
Culture preparation
E. coli O157:H7 strains F06M-0923-21 (spinach outbreak strain from California Department of Health), F07M-020-1 (Taco John's outbreak strain from California Department of Health), and avirulent strain B6914 (Dr. Pina Fratamico, Eastern Regional Research Center, USDA-ARS) were used in the study. All the strains were induced to rifampin resistance as described previously (Rice and Bonomo, 2005).
Three E. coli O157:H7 strains were grown separately in 10 mL of tryptic soy broth (Difco; Becton, Dickinson and Company, Sparks, MD) containing 100 μg/mL of rifampin (TSB-R) at 37°C. The overnight grown culture were then propagated in 20 mL each of TSB-R without Dextrose (TSB-R w/o D; pH 7.0; Difco) by inoculating a loopful of culture and incubated overnight at 37°C (control culture). Media without dextrose was used to prevent acid-adaptation due to acids produced after dextrose metabolism. Since low pH in the gastrointestinal tract of cattle is a potential place of acid-adaptation of pathogens, each strain was grown at 37°C in TSB-R w/o D with pH 5.0 (adjusted with 1 N HCl) to develop acid-adaptation response. After overnight incubation, cultures were washed twice with sterile saline and resuspended to an optical density (OD600) of approximately 0.6–0.7 (approximately 109 colony-forming unit [CFU]/mL). Equal volume of three E. coli O157:H7 cultures were mixed as inocula for the following experiments.
Inoculation of fresh dairy compost
Both control and acid-adapted cultures of E. coli O157:H7 as described above were inoculated into 2.8 kg of fresh compost mix with 50% moisture at a final concentration of approximately 107 CFU/g separately as described previously (Singh et al., 2011). The inoculated levels of the pathogens in compost were enumerated by plating serial dilutions of the compost in 0.85% saline on tryptic soy agar containing 100 μg/mL of rifampin (TSA-R; Becton, Dickinson and Company). The level of background microflora in fresh dairy compost was also enumerated by plating serially diluted compost on TSA. The finished dairy compost (Black Gold Compost Company, Oxford, FL) and 0.85% saline was also used as inoculation medium for the acid-adapted E. coli O157:H7. All the experiments that were conducted during this study have also been shown in a flow chart (Fig. 1).
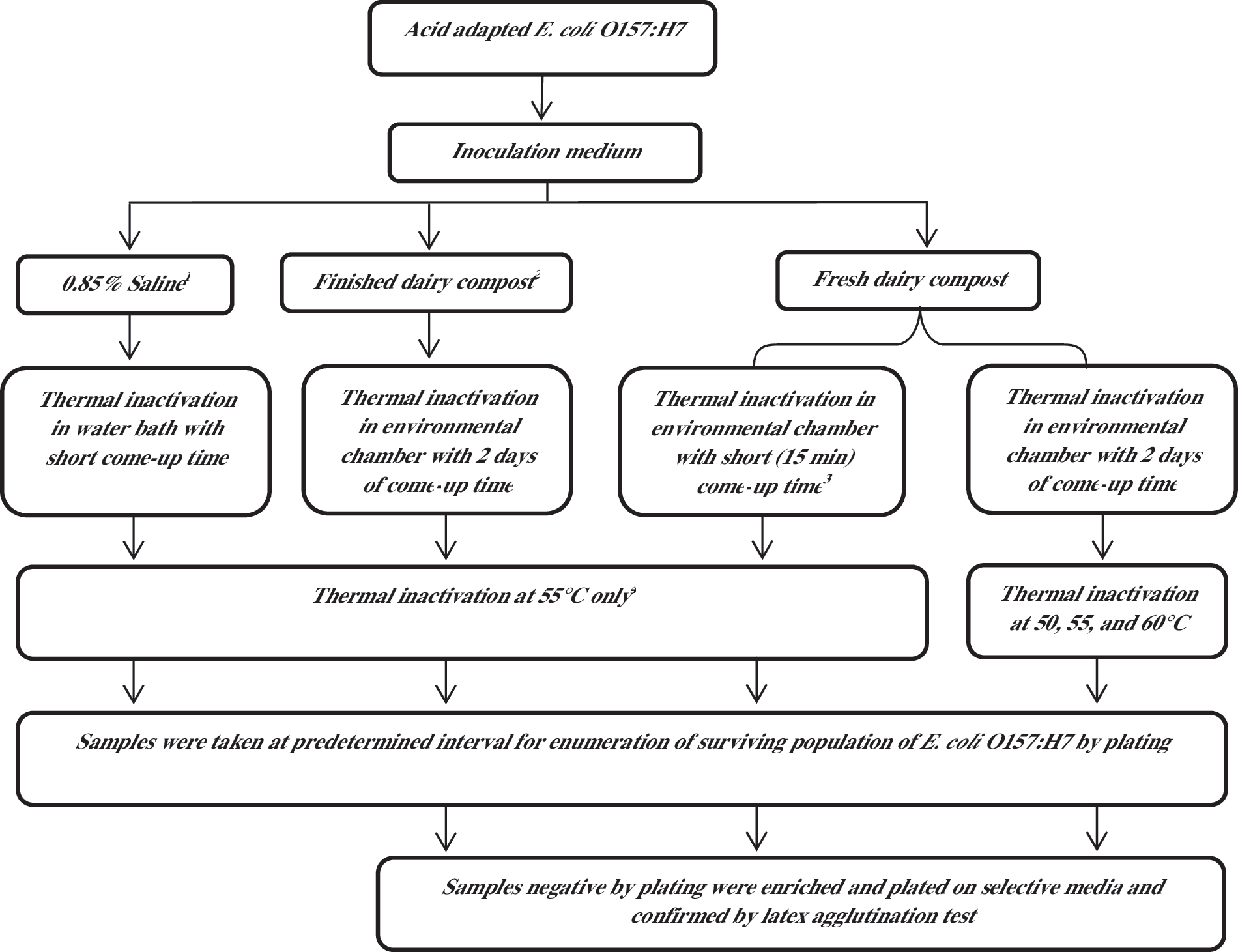
Flow chart showing complete experimental design of this study. 1Saline was used in the study to see the effect of heating medium (liquid) on thermal inactivation of acid-adapted E. coli O157:H7. 2Finished dairy compost was used to see the effect of the type of compost on thermal inactivation of acid-adapted E. coli O157:H7. 3Short come-up time study determined if loss in cross-protection was due to long come-up time of 2 days, and the experiment was done as per our previous study (Singh et al., 2010). 4Thermal inactivation at 55°C was conducted only for comparing with our actual study in fresh dairy compost.
Thermal inactivation study
The individual strains of acid-adapted and control cultures were thermally inactivated in 0.85% saline at 55°C (Fig. 1) in a circulating water bath (type 003-3332; Haake, Ober-Mörlen, Germany) and then plated on TSA-R after serial dilutions in saline.
For compost study, inoculated fresh compost (approximately 100 g) was packed in a Tyvek® pouch (size 5.25 in×10 in; SPS Medical, Rush, NY) and spread evenly into a thin layer (approximately 1 cm in depth). Tyvek® pouch is a chemically inert material made of spunbounded olefin and mylar that provides sterile packaging protection and is breathable to allow the exchange of oxygen and water. The unsealed pouches with compost were then kept in a single layer on the shelf of an environmental chamber (model no. EC2047N
The duplicate sample bags were removed at predetermined time intervals at each sampling point after the temperature of the compost inside the bag reached the target temperature and cooled immediately in an ice water bath. The compost sample was then processed as mentioned elsewhere (Singh et al., 2011). A 250-μL sample of compost homogenate was spiroplated (Autoplate® 4000; Spiral Biotech, Bethesda, MD) to increase the detection limit to 10 CFU/g.
The samples, negative for E. coli O157:H7 by plating, were pre-enriched and confirmed as reported earlier (Singh et al., 2011).
For comparing the effect of different types of compost and short come-up time (15 min) on thermal inactivation of acid-adapted E. coli O157:H7, a thermal inactivation study in commercial dairy compost with 50% moisture content and in fresh dairy compost, respectively, only at 55°C, was done as per Figure 1.
Statistical analysis
To compare the survival difference in bacterial populations with or without acid-adaptation plate count data were converted to log10 values, and subjected to analysis of variance with a test criterion (F statistic) and type I error controlled at p=0.05. The Tukey's multiple comparison procedure of the Statistical Analysis System (version 2001; SAS, Cary, NC) was used.
Results
The effect of acid-adaptation on thermal resistance of E. coli O157:H7 was studied in fresh dairy compost, finished compost, and 0.85% saline under various laboratory conditions (Fig. 1). The level of background microflora and pH in fresh dairy compost was 9.28±0.06 log CFU/g and 9.51±0.25, respectively, as compared with 6.68±0.05 log CFU/g and 8.48±0.12, respectively, in finished compost.
Cross-protection to heat due to acid-adaptation in saline
The acid-adapted cultures of all three strains survived better (p<0.05) than the control culture in saline at almost all sampling times when exposed to temperature of 55°C for up to 1 h (Table 1).
Mean bacterial population log colony-forming unit (CFU)±standard deviation (SD) with different upper case letter within column differs significantly (p<0.05) between two types of culture at the same heat exposure time for each strain.
ND, not detected by direct plating.
Thermal inactivation in fresh dairy compost
At 50°C, control and acid-adapted cultures survived for 17 and 19 days, respectively (Table 2). There were approximately 4.21 and 4.44 log reductions in control and acid-adapted cultures, respectively, when the temperature in the compost reached the set level (0 h). The decline thereafter in both control and acid-adapted cultures was gradual (Table 2). The pathogen survival was not different (p>0.05) at most of the sampling times between control and acid-adapted cultures.
Temperature inside the compost after 1 day (-24 h) was approximately 38°C, 40°C, and 43°C at 50°C, 55°C, and 60°C, respectively, in compost trials.
Mean log colony-forming unit (CFU)±standard deviation (SD) with different upper case letter differs significantly (p<0.05) between two treatments at that sampling interval.
Positive by enrichment.
Negative by enrichment.
NA, not analyzed.
At 55°C, both control and acid-adapted cultures survived for 144 h (6 days) as detected by enrichment (Table 2). The decline in the population of control and acid-adapted cultures were approximately 4.65 and 5.24 log reductions when the temperature inside the compost reached the target level (0 h). The decline in the population of E. coli O157:H7 in control and acid-adapted cultures were gradual once the temperature reached the set level. The survival differences between control and acid-adapted cultures were not different (p>0.05) at almost all the sampling times.
At higher temperature of 60°C, inactivation in the population of E. coli O157:H7 was quicker in comparison to 50°C and 55°C with populations of control and acid-adapted cultures surviving for 96 h, as detected by enrichment (Table 2). The populations of control and acid-adapted cultures were reduced by approximately 5.03 and 5.10 log CFU/g, respectively, when temperature of the compost reached set value (0 h). The differences in the survival of control and acid-adapted cultures were not different (p>0.05) at most of the sampling times.
Thermal inactivation with short come-up time
In order to evaluate the effect of long come-up time (2 days) on maintenance of acid-adaptation response on thermal inactivation of E. coli O157:H7 during composting, a trial with short come-up time (15 min) was conducted at 55°C in fresh dairy compost. The control and acid-adapted cultures survived for 6 and 2 h, respectively (Table 3). During come-up time of 0.25 h, the population of control and acid-adapted cultures were reduced approximately 0.73 and 0.69 log, respectively. Also, inactivation of acid-adapted culture was slower (p<0.05) at sampling times of 0.25 and 0.5 h than the control. However, after 1 h at 55°C, acid-adapted culture was inactivated more rapidly (p<0.05) than the control.
Mean log colony-forming unit (CFU)±standard deviation (SD) with different upper case letter differs significantly (p<0.05) between two treatments at that sampling interval.
Positive by enrichment.
Negative by enrichment.
Thermal inactivation in finished compost
To determine if compost texture, homogeneity, and microflora have any effect on thermal inactivation of acid-adapted E. coli O157:H7, a trial in finished compost was done (Fig. 1). The pathogen in the finished compost survived for only 24 h in both control and acid-adapted cultures as detected by enrichment (Table 4). The decline in both control and acid-adapted cultures was gradual within 24 h (-24 h), followed by a rapid decrease of approximately 5.53 and 5.14 log, respectively, when the temperature reached the set value (0 h). The reduction was not different (p>0.05) at most of the sampling times. Pathogen decline during the first 24 h was negligible in finished dairy compost, whereas approximately 1.44–2.39 log reduction was observed in fresh dairy compost (Table 2), which could be due to the difference in heat penetration rate and competitive microflora.
Temperature inside the compost after 1 day (-24 h) was approximately 40°C at 55°C.
Mean log colony-forming unit (CFU)±standard deviation (SD) with different upper case letter differs significantly (p<0.05) between two treatments at that sampling interval.
Positive by enrichment.
Negative by enrichment.
Moisture changes in compost during thermal inactivation study
Moisture loss from dairy compost with 2 days of come-up time at all three temperatures was approximately 35.5–39.9% when compost temperature reached the set value (0 h) for both types of treatment (Table 5). In fresh compost with short come-up time (15 min), the moisture loss was comparatively slow within 24 h of the sampling, with loss of approximately 17.3% and 18.6% in control and acid-adapted cultures, respectively. The loss of moisture in commercial finished compost was even higher than the fresh compost, with control and acid-adapted cultures losing approximately 41.6% and 42.6%, respectively, when temperature inside the compost reached 55°C.
Trial with short come-up time (15 min), initial moisture in control and acid-adapted was 51.05±0.23 and 50.71±0.56, respectively.
Last sample for the experiment when the E. coli O157 detection was negative by enrichment.
NA, no sample was available at that time for the experiment.
Discussion
Pathogen inactivation at high temperatures during composting is critical for the compost safety. However, a microbial cell has the ability to cross-protect to an unrelated stress (Rodriguez-Romo and Yousef, 2005). Therefore, microbial populations developing cross-protection to high temperature due to acid-adaptation is of practical importance for composting process.
In this study, we investigated effect of cross-protection on thermal inactivation of acid-adapted E. coli O157:H7 in saline or in compost by simulating early phase of composting process. Our results indicated that in saline, acid-adaptation resulted in extended survival (p<0.05) of E. coli O157:H7 at 55°C for 1 h; however, in fresh compost, this cross-protection was not observed at higher composting temperatures of 55°C and 60°C. Similar to these results, Singh et al. (2006) also reported lower cross-protection in acid-adapted E. coli O157:H7 to heat at 62°C and 65°C in ground beef. On the contrary, acid-adapted E. coli O157:H7 exhibited higher D-values than non-adapted cultures at 52°C in fruit juice (Ryu and Beuchat, 1998, 1999). Buchanan and Edelson (1999) also reported cross-protection in acid-adapted E. coli O157:H7 when thermally inactivated at 58°C in lab media, milk, chicken broth, and apple juice. Based on their study and findings reported elsewhere, Singh et al. (2006) concluded that acid-adaptation might increase cross-protection to heat in liquid food, but the same effect is lost in solid food matrix, which is in agreement with our results in saline and in compost matrix.
Microorganisms stress adaptation mechanism is under genetic control and activated on exposure to sublethal stresses such as heat, acid, and starvation (Chung et al., 2006). Microorganisms retain this mechanism until they return to their normal physiological state, which depends upon their growth conditions and storage after stress or may be strain dependent (Murano and Pierson, 1992; Mckellar and Knight 1999; Rowe and Kirk, 2000; Leenanon and Drake 2001). Lin et al. (1996) found that acid-adaptation in enterohemorrhagic E. coli persisted for 1 month under refrigeration conditions. In another study, thermotolerance of L. monocytogenes was lost within 1 h when stressed cells were immediately returned to the growth temperature of 35°C, suggesting that induction of stress was not long-lived (Bunning et al., 1990). In our study, we did see that acid-adaptation of E. coli O157:H7 was able to provide protection against heat in saline solution, and in fresh compost when inactivated immediately (short come-up time), but the protection was for a short period of time which lasted for 30 min. However, acid-adaptation of E. coli O157:H7 was unable to provide cross-protection against heat during composting with 2 days of come-up time suggesting loss of acid-adaptation response during long come-up time. Therefore, based on these observations, we can conclude that cross-protection due to acid-adaptation of E. coli O157:H7 in dairy compost was probably induced but was for short duration.
Induction of heat-shock in microorganism provides protection against lethal temperature (Murano and Pierson, 1992; Singh et al., 2010). In the present study, acid-adaptation does not seem to provide additional protection to the acid-adapted culture during composting with 2 days of come-up time. However, both control and acid-adapted cultures survived for long duration at all three temperatures used in the study. This longer survival of the pathogen especially when temperature rise of the composting process was gradual (2 days) could possibly be due to induction of heat-shock response (Singh et al., 2011). In the real-world composting process, temperature rise in compost heap is similar to what we have simulated in the laboratory (Shepherd et al., 2010). So, these results again underline the importance of temperature evolution during composting process and the possibilities of microorganism surviving longer due to heat-adaptation. These results also point to the fact that previous acid-adaptation of microbial communities does not affect their inactivation by heat, as those stresses are either lost during the mesophilic phase of the composting process or induction of heat-shock response may overpower any other previous stress adaptations.
Besides temperature, pathogen inactivation during composting is also affected by various other factors such as moisture content, carbon to nitrogen ratio (C:N), pH, aeration, and heap size (Sherman 2005). In the present study, moisture loss during the 2 days of come-up time would have been an additional contributing factor for stress adaptation of the pathogen during the trial, which can be supported by the fact that E. coli O157:H7 was inactivated faster in dairy compost with short come-up time. Singh et al. (2011) in their study also found that compost samples at the end of the long mesophilic phase (5 days of come-up time) lost more moisture than the compost samples with optimal mesophilic phase (2 days of come-up time) leading to prolonged survival of E. coli O157:H7 in compost with long mesophilic phase of composting.
Conclusion
Our results clearly demonstrated that in saline solution, acid-adaptation of E. coli O157:H7 provides cross-protection to thermal inactivation; however, there was no difference in the survival of control and acid-adapted pathogen in fresh dairy compost that simulated optimal mesophilic phase of the composting process. Thus, the acid-adaptation of pathogen only provides short term cross-protection in compost when they were exposed to heat immediately depending upon the type of heating medium. Based on these results, we conclude that the acid-adaptation of E. coli O157:H7 before composting plays a minimal role in providing cross-protection to thermal inactivation and was lost during mesophilic phase of composting.
Footnotes
Acknowledgments
The research was partially supported by a grant from Center for Produce Safety at University of California at Davis, and a grant from the USDA-NIFSI.
Disclosure Statement
No competing financial interests exist.