
Abstract
Vibrio parahaemolyticus strain 690 was isolated from gastroenteritis patients. Its thermal and ethanol stress responses have been reported in our previous studies. In this study, we further investigated the effects of various acid adaptation conditions including pH (5.0–6.0) and time (30–90 min) on the acid tolerance in different growth phases of V. parahaemolyticus 690. Additionally, the adaptive acid tolerance among different V. parahaemolyticus strains was compared. Results indicated that the acid tolerance of V. parahaemolyticus 690 was significantly increased after acid adaptation at pH 5.5 and 6.0 for 30–90 min. Among the various acid adaptation conditions examined, V. parahaemolyticus 690 acid-adapted at pH 5.5 for 90 min exhibited the highest acid tolerance. The acid adaptation also influenced the acid tolerance of V. parahaemolyticus 690 in different growth phases with late-exponential phase demonstrating the greatest acid tolerance response (ATR) than other phases. Additionally, the results also showed that the induction of adaptive ATR varied with different strains of V. parahaemolyticus. An increase in acid tolerance of V. parahaemolyticus was observed after prior acid adaptation in five strains (556, 690, BCRC 13023, BCRC 13025, and BCRC 12864), but not in strains 405 and BCRC 12863.
Introduction
Acid adaptation or acid tolerance response (ATR) has been reported in various Gram-negative and Gram-positive foodborne pathogens, including Escherichia coli O157:H7 (Yuk and Marshall, 2004), Salmonella typimurium (Arvizu-Medrano and Escartín, 2005), Shigella flexneri (Tetteh and Beuchat, 2003), Bacillus cereus (Browne and Dowds, 2002), and Listeria monocytogenes (Skandamis et al., 2008). Bacteria have demonstrated enhanced resistance to severe lethal acidic conditions when they were previously exposed to a mild sub-lethal acidic treatment (Foster and Hall, 1990; Hill et al., 1995). In addition, acid adaptation can also induce cross-protection against other stresses (Leyer and Johnson, 1993; Tosun and Gönül, 2003). This suggests that ATR may be able to contribute to the survival ability or disease-causing capability of pathogens, and it should be considered for microbial food safety risk assessments.
V. parahaemolyticus is a moderately halophilic bacterium that is indigenous to the marine environments worldwide (Beuchat, 1982). It is commonly isolated from a variety of seafoods and can cause acute human gastroenteritis associated with the consumption of contaminated seafood (Liston, 1990; Feldhusen, 2000; Su and Liu, 2007). The incubation period for V. parahaemolyticus infections range from 4 to 96 h. The most common symptoms of gastroenteritis caused by this pathogen include diarrhea, abdominal cramps, nausea, vomiting, headache, fever, and chills (Daniels et al., 2000; Nair et al., 2007). This pathogenic microorganism has been recognized as a major cause of foodborne illness in China, Japan, Taiwan, and other Asian coastal regions, where people often consume raw or minimally processed seafood (Wong et al., 2000; Bhuiyan et al., 2002; Hara-Kudo et al., 2003; Wang et al., 2007). V. parahaemolyticus accounted for 63.7% of the total bacterial foodborne outbreaks in Taiwan from 1981 to 2010 (Anonymous, 2011).
The acid resistance of V. parahaemolyticus is an important factor for its ability to withstand acidic challenges in the food systems. The adaptive ATR has also been characterized in other Vibrio spp., including V. cholerae and V. vulnificus (Alvarez et al., 2003; Bang and Drake, 2005). In addition, studies have revealed that the ATR of pathogenic bacteria can be influenced by the type of medium and acidulant, adaptation and challenge conditions, bacterial strain, and growth phase (Bearson et al., 1997; Foster, 1999; Deng et al., 1999; Tetteh and Beuchat, 2001; Jobin et al., 2002; Faleiro et al., 2003; Arvizu-Medrano and Escartín, 2005; Bang and Drake, 2005; Skandamis et al., 2008; Álvarez-Ordóñez et al., 2009). These factors might also be involved in acid tolerance in V. parahaemolyticus. However, related information concerning this pathogen is still limited, and thus we elected to conduct further studies.
We have found that heat and ethanol adaptations increased the thermal and ethanol tolerances of V. parahaemolyticus strain 690, but decreased its resistance to acid and high-salt stresses. These treatments also altered its protein expression, fatty acid composition, and superoxide dismutase and catalase activities (Chang et al., 2004; Chiang et al., 2005, 2006, 2008a,b; Chiang and Chou, 2008, 2009). According to these findings, the objective of this study was to further investigate the effects of various acid adaptation conditions on the acid tolerance in different growth phases of V. parahaemolyticus 690. Additionally, the adaptive acid tolerance among different V. parahaemolyticus strains was compared.
Methods
Microorganisms
Seven strains of V. parahaemolyticus were used as the test organisms. V. parahaemolyticus 405, 556, and 690, originally isolated from clinical samples of gastroenteritis patients, were obtained from Professor H.C. Wong, Department of Microbiology, Soochow University (Taipei, Taiwan). V. parahaemolyticus BCRC 12863 (American Type Culture Collection [ATCC] 17803) and BCRC 12864 (ATCC 27519), involved in food poisoning outbreaks in Japan and the United States, respectively, and BCRC 13023 and BCRC 13025, associated with food poisoning outbreaks in Taiwan, were purchased from Bioresource Collection and Research Center (BCRC), Food Industry Research and Development Institute (FIRDI; Hsinchu, Taiwan). The test organisms were activated by two successive transfers in tryptic soy broth (TSB; Acumedia Manufactures, Lansing, MI) supplemented with 3% NaCl (TSB-3% NaCl) at 37°C for 6 h; these activated cultures served as the inocula of experiment.
Growth study at different pH values
V. parahaemolyticus 690 was examined for growth at different pH values. First, 50 mL of TSB-3.0% NaCl was acidified with 10.0 N HCl to pH 4.0, 4.5, 5.0, 5.5, 6.0, 6.5, and 7.0, respectively. Then, 0.1 mL of the above activated culture was inoculated into the acidified TSB-3.0% NaCl at an initial population of approximately 106 colony-forming unit (CFU)/mL and incubated at 37°C for 6 h. The viability of the test organism was determined every 1 h during the incubation period. Among these various pH values, pH 5.0, 5.5, and 6.0 were chosen for acid adaptation, and pH 4.5 was designated for subsequent acid challenges.
Measurement of acid tolerance during the growth cycle
The method stated in Davis et al. (1996) was adopted with minor modifications to measure the acid tolerance of V. parahaemolyticus 690 during the growth cycle. 0.1 mL of the activated culture was inoculated into 50 mL of TSB-3.0% NaCl at an initial population of approximately 105 CFU/mL and incubated at 37°C for 6 h. Samples were taken to determine the population growth every 30 min during the incubation period. Simultaneously, their viability after an acid challenge at pH 4.5 for 4 h was also examined. The cultures cultivated at 37°C for 2.5, 4.0, and 5.5 h were assumed as the mid-exponential, late-exponential, and stationary phase cultures, respectively.
Acid adaptation treatment
Acid adaptation of V. parahaemolyticus was performed according to the method of Bang and Drake (2005) with minor changes. 50 mL of the mid-exponential, late-exponential, or stationary phase culture was centrifuged (3,000×g, 10 min) and washed twice with phosphate-buffered saline (PBS) containing 3% NaCl (PBS-3% NaCl, 0.1M Na2HPO4/NaH2PO4, pH 7.0). The acid-adapted cells were prepared by suspending in 50 mL of acidified TSB-3% NaCl (pH 5.0, 5.5, and 6.0) and held at 37°C for 30, 60, and 90 min, respectively. The non-adapted cells were prepared by suspending in TSB-3% NaCl without acidification and incubation.
Determination of adaptive acid tolerance at pH 4.5
Late-exponential phase cells of V. parahaemolyticus were used to determine their acid tolerance in addition to the growth phase test. Firstly, the study investigated the effect of acid adaptation pH and time on the acid tolerance of V. parahaemolyticus 690. 1.0 mL of the acid-adapted (pH 5.0–6.0, 30–90 min) or non-adapted late-exponential phase cells was inoculated into 50 mL of acidified PBS-3% NaCl (pH 4.5) at an initial population of approximately 106 CFU/mL. The samples were incubated at room temperature (approximately 25°C) for 4 h. The viability of the test organism was determined every 40 min during the incubation period. According to the result of this experiment, an adaptive procedure at pH 5.5 for 90 min was used for acid adaptation of V. parahaemolyticus in the following trials.
Secondly, the study examined the effect of growth phase on the adaptive acid tolerance of V. parahaemolyticus 690. 1.0 mL of the acid-adapted (pH 5.5, 90 min) or non-adapted cells in mid-exponential, late-exponential, or stationary phase was inoculated into 50 mL of acidified PBS-3% NaCl (pH 4.5) at an initial population of approximately 106 CFU/mL and incubated at room temperature. The viability of the test organism was determined after exposure at pH 4.5 for 4 h.
Finally, to evaluate the adaptive acid tolerance among V. parahaemolyticus strains, 1.0 mL of the acid-adapted (pH 5.5, 90 min) or non-adapted late-exponential phase cells of test strains was inoculated into 50 mL of acidified PBS-3% NaCl (pH 4.5) at an initial population of approximately 106 CFU/mL and incubated at room temperature for a period of 4 h. At the end of the exposure period, the viability of each test strain was determined.
Enumeration of viable V. parahaemolyticus
V. parahaemolyticus samples were serially diluted in PBS-3.0% NaCl and pour plated (1 mL) with tryptic soy agar (TSA; Acumedia Manufactures, Lansing, MI) supplemented with 3% NaCl (TSA-3% NaCl). The viable colonies were enumerated after 18 h of incubation at 37°C.
Statistical analysis
All trials were repeated three separate times, and the results were expressed as mean values±standard deviations. Experimental data were analyzed by analysis of variance with SAS statistical software (SAS Institute, Cary, NC). The difference between non-adapted and acid-adapted groups was compared by unpaired two-tailed Student's t-test. The differences between acid adaptation condition, growth phase, and strain groups were compared by Duncan's multiple range test, and the statistically significant difference was set at p<0.05.
Results and Discussion
Growth of V. parahaemolyticus 690 at different pH values
In this study, the growth of V. parahaemolyticus 690 in TSB-3% NaCl at different pH values was examined. As shown in Figure 1, the growth of V. parahaemolyticus 690 was affected by the pH of media. The population of V. parahaemolyticus 690 increased with the incubation time when pH was 5.5 or greater. Contrarily, when pH was lower than 5.0, the cell counts were decreased with the incubation time. V. parahaemolyticus has been reported to grow from 5.0 to 11, and its optimum growth pH value is 7.5–8.0 (Joseph et al., 1982). Yeung and Boor (2004) reported that none of the test V. parahaemolyticus strains was able to grow after 1 h of exposure at pH 4.2 and 3.0. Based on our results and these reports, pH 5.0, 5.5, and 6.0 were selected as the conditions for acid adaptation. pH 4.5 was used as acid challenge condition in the subsequent studies.
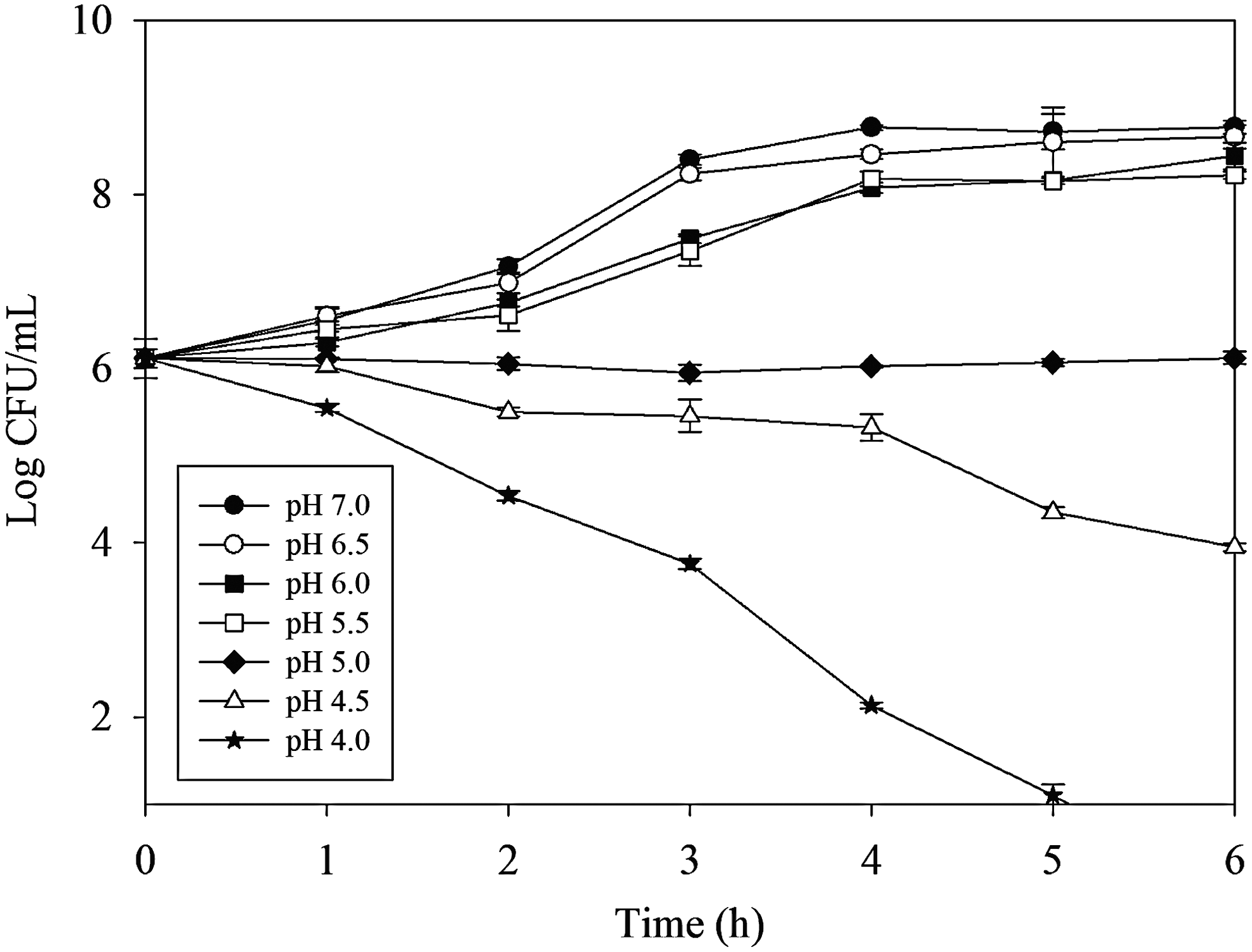
Growth of Vibrio parahaemolyticus 690 in tryptic soy broth (TSB)–3% NaCl at different pH values.
Acid tolerance of V. parahaemolyticus 690 during the growth cycle
In this study, the survival of V. parahaemolyticus 690 after exposure at pH 4.5 for 4 h was examined during the growth cycle. As shown in Figure 2, starting from 0 h of cultivation, the survival of V. parahaemolyticus 690 gradually decreased with the incubation time. A minimum survival of only 0.06% was observed at 3.5 h of cultivation in mid-exponential phase. Nevertheless, the survival rose again when the culture further cultivated to reach late-exponential and stationary phases. V. parahaemolyticus 690 grown in stationary phase showed a maximum survival of 8.01% at 6 h of cultivation. It was noted that the mid-exponential phase cells were most susceptible to acid, while the stationary phase cells exhibited the highest acid tolerance. Our findings were parallel to those of Koo and Jahncke (2006), who indicated that V. parahaemolyticus TX-2103 and V. vulnificus had the least acid tolerance in mid-log phase, while displaying the most acid tolerance in late-log and stationary phases. The phenomena might involve the induction of RpoS (σs) encoded by rpoS, which has been identified in V. parahaemolyticus (Vasudevan et al., 2006; Whitaker et al., 2010). RpoS is a sigma subunit of RNA polymerase, which is an important regulator for bacterial ATR and could be induced during the stationary growth phase (Lee et al., 1995; Bearson et al., 1996; Cheville et al., 1996; Hengge-Aronis, 2000). The occurrence of growth-phase–dependent acid tolerance has also been observed in other pathogens (Arnold and Kaspar, 1995; Benjamin and Datta 1995; Davis et al., 1996; Gorden and Small, 1993; Hill et al., 1995; Browne and Dowds, 2002; Jobin et al., 2002).

Acid tolerance of Vibrio parahaemolyticus 690 during the growth cycle. Solid circle, cell population; open circle, surviving percentage was determined after exposure to pH 4.5 for 4 h and calculated by dividing the final survival population by the initial population which corresponds to 100%.
Effect of acid adaptation condition on the adaptive acid tolerance of V. parahaemolyticus 690
Figure 3 illustrates the effect of acid adaptation time on the survival of V. parahaemolyticus 690 at pH 4.5. After 4 h of exposure, the survival between acid-adapted cells at pH 5.0 for 30, 60, and 90 min (0.22–0.42%) and non-adapted cells (0.35%) was not significantly different (p>0.05; Fig. 3A). However, when acid-adapted at pH 5.5 and 6.0, the survival of acid-adapted V. parahaemolyticus 690 generally increased as the acid adaptation time extended (Fig. 3B). A significant increasing trend (p<0.05) in the survival of V. parahaemolyticus 690 was also found when acid-adapted at pH 6.0 for 30 min extended to 90 min (Fig. 3C). The better survival abilities were also observed in acid-adapted V. vulnificus (Bang and Drake, 2005), E. coli O157:H7 (Cheng et al., 2003), and B. cereus (Chen et al., 2009) compared to their non-adapted cells. A possible explanation might be that the acid-adapted bacteria could maintain their intracellular pH (pHi) at a higher level than non-adapted cells (Browne and Dowds, 2002; Thomassin et al., 2006). Besides, synthesis of the specific acid-adaptive or acid shock proteins (ASPs) in acid-adapted cells might be another important factor to prevent or repair damaged macromolecules and promote bacterial survival in subsequent extreme acid stress (Foster, 1991, 1993; Bearson et al., 1997; Tetteh and Beuchat, 2003). Zhao and Houry (2010) also identified the acid-stress-response regulatory proteins, chaperones, and decarboxylases that allow the bacteria to respond to the external acid environment.
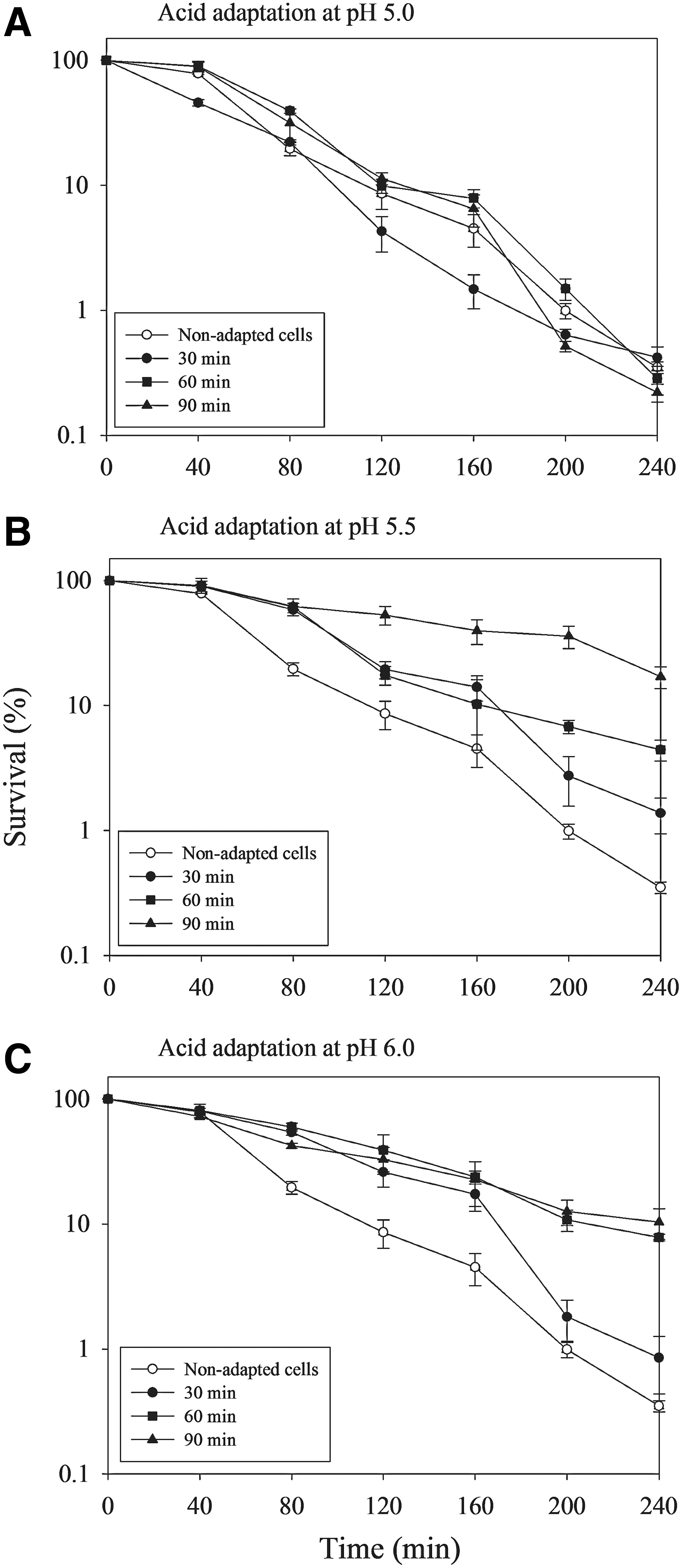
Effect of acid adaptation time on the survival of Vibrio parahaemolyticus 690 at pH 4.5. The initial populations of non-adapted and acid-adapted cells were approximately 106 colony-forming unit (CFU)/mL. Surviving percentage was obtained by dividing the survival population by the initial population, which corresponds to 100%. Data were expressed as mean±standard deviations from the three separate experiments.
Figure 4 summarizes the effect of acid adaptation pH on the survival of V. parahaemolyticus 690 at pH 4.5. When duration of acid adaptation time was similarly performed for 30, 60, or 90 min, the survival of acid-adapted V. parahaemolyticus 690 at pH 5.5 or 6.0 was higher than those of non-adapted cells and acid-adapted cells at pH 5.0 (Fig. 4A–C). Under the same duration of acid adaptation time, acid adaptation at pH 5.5 and 6.0 improved the survival of V. parahaemolyticus 690, but this improvement was not observed at pH 5.0 (Fig. 4C). This finding is different from the report of Wong et al. (1998), which described that adaptation at pH 5.0 was more effective than at pH 5.5 and 5.8 for inducing acid tolerance of V. parahaemolyticus ST550. It is speculated that different strains, possessing different sensitivities to pH, result in the diverse consequences. This study revealed that adaptive acid tolerance of V. parahaemolyticus 690 was influenced by acid adaptation conditions. Among the various acid adaptation conditions examined, cells of V. parahaemolyticus 690 acid-adapted at pH 5.5 for 90 min possessed the greatest acid tolerance, with the highest survival of 17.03%. Adaptation in a mild acid treatment increased the bacterial tolerance to a severe acid stress for V. cholerae, V. parahaemolyticus and V. vulnificus, which was also reported by Koga et al. (1999), Alvarez et al. (2003), Yeung and Boor (2004), and Koo and Jahncke (2006). According to the above findings, an acid adaptation treatment at pH 5.5 for 90 min was selected for testing the adaptive acid tolerance of V. parahaemolyticus in the subsequent studies.

Effect of acid adaptation pH on the survival of Vibrio parahaemolyticus 690 at pH 4.5. The initial populations of non-adapted and acid-adapted cells were approximately 106 colony-forming unit (CFU)/mL. Surviving percentage was obtained by dividing the survival population by the initial population, which corresponds to 100%. Data were expressed as mean±standard deviations from the three separate experiments.
Effect of growth phase on the adaptive acid tolerance of V. parahaemolyticus 690
According to the results shown in Figure 2, cells of V. parahaemolyticus 690 in mid-exponential, late-exponential, and stationary phases were further subjected to acid adaptation at pH 5.5 for 90 min, and their survival under acid challenge was determined. Table 1 shows the survival of non-adapted and acid-adapted V. parahaemolyticus 690 in various growth phases after exposure at pH 4.5 for 4 h. The survival of acid-adapted V. parahaemolyticus 690 in mid-exponential, late-exponential, and stationary phases was approximately 6.4-, 38.6-, and 2.0-fold that of non-adapted V. parahaemolyticus 690 in the same growth phase, respectively. The acid-adapted late-exponential phase cells revealed a higher extent of increased acid tolerance compared to the acid-adapted mid-exponential and stationary phase cells. The results demonstrated that adaptive acid tolerance of V. parahaemolyticus 690 varied with the growth phase of cultures. The ATR phenomenon has been found to be induced during the exponential and stationary growth phases (Hill et al., 1995; Lee et al., 1995; Davis et al., 1996; Bearson et al., 1997). In this study, the most pronounced adaptive ATR of V. parahaemolyticus 690 was noted in late-exponential growth phase. These findings are dissimilar to the results of Yeung and Boor (2004). They reported that the survival of stationary V. parahaemolyticus strains, previously acid-adapted at pH 5.5, was decreased after exposure at pH 3.6. The diverse results may be due to the differences in bacterial strain and pH condition of acid challenge used. Moreover, our previous studies also indicated that V. parahaemolyticus 690 demonstrated the greater heat and ethanol adaptation responses in late-exponential phase than in other growth phases (Chiang and Chou, 2009). Therefore, we further investigated the adaptive acid tolerance among different V. parahaemolyticus strains during late-exponential phase.
The initial populations of V. parahaemolyticus were ca. 106 CFU/mL. Surviving percentage was obtained by dividing the final survival population by the initial population which corresponds to 100%.
Values in the same column with different upper case letters (A, B) are significantly different (p<0.05). Values in the same row with different lower case letters (x, y) are significantly different (p<0.05).
Effect of bacterial strain on the adaptive acid tolerance of V. parahaemolyticus
To compare the adaptive acid tolerance among V. parahaemolyticus strains, seven strains of V. parahaemolyticus in late-exponential phase were acid-adapted at pH 5.5 for 90 min, respectively. The survival of non-adapted and acid-adapted V. parahaemolyticus strains after exposure at pH 4.5 for 4 h was shown in Table 2. Among seven V. parahaemolyticus strains, BCRC 12864 demonstrated the greatest acid tolerance, while BCRC 13023 was most susceptible to acid, at the end of the exposure at pH 4.5. The survival of acid-adapted V. parahaemolyticus was significantly (p<0.05) higher than that of non-adapted V. parahaemolyticus in five strains, including 556, 690, BCRC 12864, BCRC 13023, and BCRC 13025. Nonetheless, the survival between the acid-adapted and non-adapted V. parahaemolyticus was similar in two strains, 405 and BCRC 12863. The results showed that the adaptive acid tolerance of V. parahaemolyticus varied among the seven test strains. In addition, strain 690 exhibited the greatest extent of increased acid tolerance, followed by BCRC13023, BCRC 13025, 556, and BCRC 12864, while strains 405 and BCRC 12863 did not enhance their acid tolerance, after prior acid adaptation. Considerable differences in adaptive ATR among V. parahaemolyticus strains in our results are consistent with several studies. Bang and Drake (2005) found that acid-adapted V. vulnificus had various adaptive acid tolerances between bacterial strains. Cheng and Chou (2001) and Cheng et al. (2003) indicated that the survival of acid-adapted E. coli O157:H7 during exposure to acid conditions varied with the strains of this pathogen. Furthermore, an apparent variation of ATR was also observed in different S. typhimurium and L. monocytogenes isolates, respectively (Faleiro et al., 2003; Berk et al., 2005; Adrião et al., 2008).
The initial populations of V. parahaemolyticus were ca. 106 CFU/mL. Surviving percentage was obtained by dividing the final survival population by the initial population which corresponds to 100%.
Values in the same column with different upper case letters (A, B, C) are significantly different (p<0.05). Values in the same row with different lower case letters (x, y) are significantly different (p<0.05).
Conclusion
The results of this study showed that the induction of adaptive ATR in V. parahaemolyticus was dependent on acid adaptation condition, growth phase, and bacterial strain. Among the test strains, V. parahaemolyticus 690 induced the most pronounced ATR in late-exponential phase after acid adaptation at pH 5.5 for 90 min. These findings are critical to the understanding of the presence and survival of V. parahaemolyticus in the acidic conditions of food processing and preservation that may pose a potential greater threat to food safety. Further studies are underway to investigate the protein expression profile in V. parahaemolyticus strains to know the regulatory roles of these proteins in adaptive ATR of this pathogen.
Footnotes
Acknowledgments
This research was funded by the project of National Science Council, Executive Yuan (Taipei, Taiwan; grant NSC 99-2313-B-424-001-MY3).
Disclosure Statement
No competing financial interests exist.