
Abstract
Tetrandrine (TET) is a bis-benzylisoquinoline alkaloid derived from the radix of Stephania tetrandra S. Moore. TET performs a wide spectrum of biological activities. The radix of S. tetrandrae has been used traditionally in Asia, including Korea, to treat congestive circulatory disorders and inflammatory diseases. The aim of this study was to examine the mechanism of antibacterial activity of tetrandrine against Staphylococcus aureus. The mechanism was investigated by studying the effects of TET in combination with detergent or membrane potential un-couplers. In addition, the direct involvement of peptidoglycan (PGN) was assessed in titration assays. TET activity against S. aureus was 125–250 μg/mL, and the minimum inhibitory concentration (MIC) of the two reference strains was 250 μg/mL. The OD600 of each suspension treated with a combination of ethylenediaminetetraacetic acid (EDTA), tris(hydroxymethyl) aminomethane (TRIS), and Triton X-100 (TX) with TET (0.25×MIC) had been reduced from 43% to 96%. Additional structure-function studies on the antibacterialactivity of TET in combination with other agents may lead to the discovery of more effective antibacterial agents.
Introduction
S. aureus (family, Micrococcaceae) is a Gram-positive coccus that grows in clusters and is a part of the human indigenous microflora (Lowy, 1998; Chomnawang et al., 2009). The transmission of S. aureus causes both endemic and epidemic diseases. Many strains of S. aureus are developing resistance against available antibacterial agents, leading to complications in treating infections related to skin, soft-tissue, respiratory tissue, bone, joint, and endovascular system disorders (Herwaldt, 1999). We hypothesized that the direct effects of TET on the bacterial cell wall may be responsible for the synergy between many agents and TET against S. aureus (Zhao et al., 2001).
Therefore, we investigated the in vitro activities of TET separately or TET in combination with the bacterial membrane-binding agents ethylenediaminetetraacetic acid (EDTA), tris(hydroxymethyl)aminomethane (TRIS), and Triton X-100 (TX), and the ABC transporter-inhibiting agents sodium azide (NaN3) and N,N′-dicyclohexylcarbodiimide (DCCD). In addition, we sought ways to overcome discrepancies in TET activity (Jung et al., 2008). We also investigated the effects of adding peptidoglycan (PGN) derived from S. aureus into Mueller-Hinton broth (MHB) that contained TET alone (Zhao et al., 2001).
In this study, we aimed to gain insights into the antibacterial mechanism of TET against S. aureus by evaluating the relationship between membrane permeabilization and the combined effects of TET and PGN.
Methods
TET and other reagents
TET, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), DCCD, NaN3, TX, EDTA, and purified lipopolysaccharide (LPS; obtained from Escherichia coli O26:B6) were all purchased from Sigma-Aldrich Co. (St. Louis, MO). Purified PGN (obtained from S. aureus) and TRIS were purchased from Fluka Chemika (Buchs, Switzerland) and Amresco (Solon, OH), respectively.
S. aureus strains and culture medium
MHB and Muller-Hinton agar (MHA) were purchased from Difco Laboratories (Baltimore, MD). In this study, we used S. aureus of American Type Culture Collection (ATCC; Manassas, VA) and Culture Collection of Antimicrobial Resistant Microbes (CCARM; Seoul, Korea) (ATCC25923, ATCC33591, CCARM 3090, CCARM 3091, CCARM 3095, and CCARM 3102). The ATCC and CCARM strains were purchased commercially, and CCARM clinical isolates were obtained from four different patients at Asan Medical Center (Seoul, Korea). Prior to use, all bacterial isolates were stored in 30% glycerol at −70°C. All the strains were maintained on MHA or nutrient agar plates, and the antibacterial assays were performed using MHB. Bacterial growth was monitored by measuring the turbidity of the culture at an optical density (OD) of 600 nm (Zhao et al., 2001; Jung et al., 2008).
Minimum inhibitory concentration assay
The minimum inhibitory concentration (MIC) assay was performed according to the method recommended by the Clinical and Laboratory Standards Institute (CLSI, 2000) guidelines and by Saiful et al. (2008) and Mohtar et al. (2009). The MIC values for TET against S. aureus were determined using the double-broth microdilution method in 96-well microtiter plates. Serial twofold dilutions of TET dissolved in DMSO (v/v diluted with MHB) or antibacterial agents dissolved in MHB were prepared in successive wells of 96-well microtiter plates. Then, a microbial suspension (108 colony forming units [CFU]/mL) cultured overnight was added to the wells, and the plates were incubated at 37°C for 24 h. For the serial dilutions, a 50-μL aliquot of the antibacterial agents is singular, or the TET solution was added to well A in a 96-well plate and diluted using either MHB or 15% DMSO in MHB to a final volume of 100 μL. In addition, 50 μL of MHB or 15% DMSO in MHB was added in wells B–H. The wells were serially diluted from well A through well H. The final concentration of DMSO was 5% (v/v) after dilution, which did not inhibit bacterial growth (Mohtar et al., 2009). The 5% DMSO did not show any inhibitory effects. For further confirmation, 20 μL (1 mg/mL) of MTT reagent was added to the suspension in selected wells after 24-h incubation, which was followed by aerobic incubation for 30 min at 37°C. Wells appearing yellowish-clear in color indicated inhibition of microbial growth, and wells dark blue in color indicated the absence of inhibition. MIC is defined as the lowest concentration that produces no visible growth (i.e., absence of turbidity and or precipitation), as observed with the naked eye (Saifu et al., 2008; Mohtar et al., 2009).
Antibacterial activity with detergents and ATPase inhibitors
To elucidate whether the antibacterial activity elicited by TET was associated with altered membrane permeability, the antibacterial susceptibility of TET was examined in the presence of detergents or ATPase inhibitors. To increase membrane permeability, TET was added to bacterial cells at a concentration of 0.25×MIC (1/4 MIC) and sub-inhibitory concentration (SIC) of TET was determined by a combination assay with the following other therapeutic reagents: 0.009% TX, 31.25 mM Tris, and 0.19 mM EDTA. NaN3 and DCCD were used as ATPase inhibitors (Linnett et al., 1979). The antibacterial activity of TET including 0.00125% NaN3 and 50 μM DCCD was tested as described above.
TET and PGN combination assay
TET and PGN combination assays were performed in a manner similar to the combination assay, with modifications: the 0.5 McFarland standardized overnight cultures were inoculated into MHB containing a series of PGN concentration. Serial dilutions of PGN (0–62.5 μg/mL) were mixed with TET at half the MIC value (125 μg/mL). LPS was used as the control (Zhao et al., 2001).
Statistical analysis
All experiments were performed three times or more, and the data presented were an average of those experiments. The data were presented as mean±SD values for the indicated number of separate experiments. In general, statistical significance was set at 0.05. The data were analyzed using SPSS 17.0 for Windows (SPSS Inc., Chicago, IL).
Results
Antibacterial susceptibility
The results of the MIC assays are shown in Table 1. Similar MIC values for TET were recorded in all test isolates. The MIC values of TET against all strains were 125–250 μg/mL and that of AMP ranged from 0.9 to >250 μg/mL. The MIC value of OXA was 1.9 μg/mL against ATCC 25923 strain, and 125 to >250 μg/mL against the other strains. Most S. aureus strains were significantly resistant to AMP and OXA. In addition, the MIC value of ATCC 25923 was measured at OD600 nm (Fig. 1). The results of the OD600 value indicate that bacterial growth does not significantly change in 1/2 or less MIC treatment compared to normal media (or no treatment or a negative control). Figure 1 also shows the results obtained when S. aureus (ATCC 25923) was treated with TET.

Antibacterial activity of tetrandrine (TET) against Staphylococcus aureus (American Type Culture Collection [ATCC] 25923).
ATCC, American Type Culture Collection; CCARM, Culture Collection of Antimicrobial Resistant Microbes.
Antibacterial activity with detergents or ATPase inhibitors
To investigate the effects of detergent-induced membrane permeability on the activity of TET, the antibacterial activity of TET in S. aureus strains with increased membrane permeability was examined using previously determined concentrations (0.19 mM EDTA, 31.25 mM Tris, and 0.009% TX). Tris and TX are membrane-permeabilizing agents that increase membrane permeability in organisms by binding LPS or PGN (Leive, 1965; Irvin et al., 1981; Wang et al., 2007). We noted modest reductions in the OD600 value of TET-treated suspensions (Fig. 2A–C). Compared to the OD600 value of TET alone (0.25×MIC), the OD600 values of the suspensions treated with the combination of EDTA, Tris, and TX with TET were reduced to 56%, 96%, and 44%, respectively. We investigated bacterial viability in the presence of TET, with 0.00125% NaN3 as a metabolic inhibitor, which can decrease ATP levels by disrupting electrochemical proton gradients in a bacterial environment (Swallow et al., 1990; Jernaes et al., 1994; Goncalves et al., 1999). However, TET in combination with either NaN3 or DCCD did not reduce the viability (Fig. 2D,E).

The impact of membrane-permeabilizing agent and ATPase-inhibitor agent on Staphylococcus aureus susceptibility to tetrandrine (TET). Bacterial viability was determined with a spectrophotometer (optical density at 600 nm) after 24-h incubation with TET at 0.25×minimum inhibitory concentration (MIC; 1/4 MIC) and the indicated concentration in American Type Culture Collection (ATCC) 25923.
Direct binding of TET with peptidoglycan and lipopolysaccharide
TET may bind directly to the S. aureus cell wall, thereby interfering with its integrity. The direct binding of TET with PGN was confirmed by adding PGN or LPS (0–62.5 μg/mL) derived from S. aureus into MHB containing TET alone (125 μg/mL). At a concentration of 62.5 μg/mL, PGN inhibited the antibacterial activity of TET (Fig. 3A). At a concentration of 62.5 μg/mL, LPS did not show any inhibitory effects (Fig. 3B). PGN or LPS alone showed no effect on the antibacterial effect of TET. These results suggest that TET binds directly with PGN.
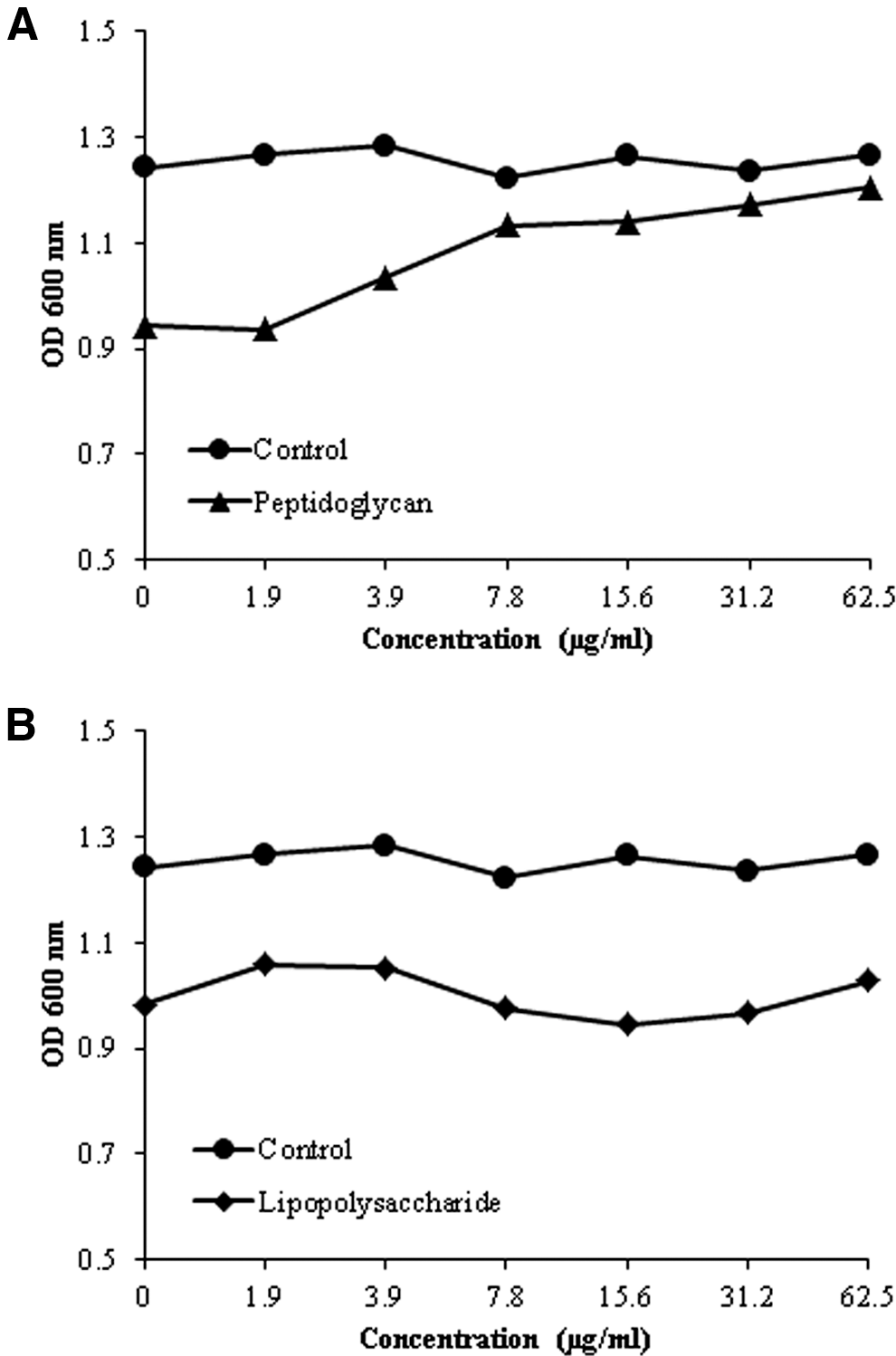
Direct binding of tetrandrine (TET) to peptidoglycan (PGN) or lipopolysaccharide (LPS) in the cell wall. Staphylococcus aureus–derived PGN was added to Mueller-Hinton broth (MHB) that contained TET alone
Discussion
S. aureus is an important human pathogen; it causes life-threatening systemic infections such as pneumonia, septicemia, endocarditis, and osteomyelitis (Lee et al., 2008). Furthermore, S. aureus can easily spread from person to person and has been detected in the noses of approximately 40–50% of healthy people and is also commonly found on the skin (Marraro et al., 1975; Sanford et al., 1994; Lowy, 1998). TET, a bis-benzylisoquinoline alkaloid derived from the root of S. tetrandra, is known to induce antibacterial activity against S. aureus (Lee et al., 2011). However, the precise mechanism by which TET inhibits the growth of S. aureus has not been elucidated.
The MIC assay is considered to be the standard method for determining the susceptibility of various microorganisms to antibacterial agents. Our in vitro results show that the MIC values of TET, AMP, and OXA against S. aureus are within the ranges that have been reported in previous studies (Dzink et al., 1985; Kwon et al., 2007; Choi et al., 2010). Antibacterial agents such as β-lactams inhibit bacterial growth by using a number of complex mechanisms, including the inhibition of cell wall synthesis (Al-Habib et al., 2010). In this study, TET was used in combination with detergents (EDTA, Tris, and TX) responsible for increasing the membrane permeability of bacterial strain. After 24-h incubation, S. aureus viability or the OD value decreased by 96% (TET concentration, 0.25×MIC) when exposed to a combination of TET and Tris, in comparison with the value observed when exposed to TET alone and the control value. In contrast, TET in combination with ATPase inhibitors (NaN3 and DCCD) had no effect on the OD values (Fig. 2D,E); this finding may indicate that the primary mechanism is not via ATPase inhibitors. These results are consistent with those of a previous study, which reported that S. aureus showed slightly greater resistance to NaN3 than Gram-negative bacteria (Lichstein et al., 1944). However, the effects of NaN3 and DCCD are broader than just ABC transporters. NaN3 is an inhibitor of cytochrome c oxidase, and DCCD inhibit proton-driven ATPases via distinct mechanisms.
The PGN content in the cell wall of Gram-negative bacteria is less than 10%, whereas it ranges from 5% to 95% in the cell wall of Gram-positive bacteria (Farca et al., 1994). This difference may account for the increased resistance of Gram-positive bacteria on exposure to EDTA-TET, Tris-TET, and TX-TET. Nevertheless, the lesser potential for antibacterial activity against S. aureus strains can be further explained. Cells are protected from lysis in low osmotic environments because their cell walls have high tensile strength. Any damage to the cell wall certainly will decrease cell tolerance to high ionic strength and low osmotic pressure (Zhao et al., 2001). The cell wall in Gram-positive bacteria is composed of 30–50 PGN sheets outside the cell membrane, and it plays an essential role not only in osmotic protection but also in cell division (Engels et al., 2011). The cell wall also serves as a primer for its own biosynthesis. In Gram-negative bacteria, however, the PGN layer is thin (one or two sheets) and is overlaid by an outer membrane composed mainly of LPS (Zhao et al., 2001). The different susceptibilities to TET may be explained primarily by the structural differences between Gram-positive and Gram-negative bacteria, and the low affinity between TET and LPS. These results indicate that TET damages the cell wall.
TET may synergize with β-lactam activity because both TET and β-lactams directly or indirectly attack the same target: PGN in the cell wall (Zhao et al., 2001). TET-induced damage of the bacterial cell wall along with potential interference of cell wall biosynthesis via its direct binding with PGN may underlie the major synergy against S. aureus (Zhao et al., 2001). The additive or neutral effects of TET in combination with other agents strongly support this explanation.
Therefore, TET may be a good antibacterial drug candidate for clinical development. The results of the present study appear promising and may increase the use of natural products as drugs.
Footnotes
Acknowledgments
This study was supported by the 2012 Post-Doctoral Fellowship Program of National Institute of Horticultural & Herbal Science, Rural Development Administration, Republic of Korea.
Disclosure Statement
No competing financial interests exist.