
Abstract
Salmonella is an international foodborne pathogen widely disseminated in seawater that regularly causes large outbreaks of food poisoning. In this study, we have investigated the effect of starvation on the ability of Salmonella enterica serovar Typhimurium cells to adhere to polystyrene microplate and Hep2 cells in seawater microcosms after incubation for 3 years. Cell surface hydrophobicity was evaluated. Effect of stress on fatty acids composition was also established. Our results showed that after incubation in seawater, the ability of starved cells to adhere to polystyrene microplate was decreased significantly. However, the adhesion values to Hep2 cells have increased. In addition, cells surface hydrophobicity was decreased. The fatty acids composition of starved cells was modified.
Introduction
E
Generally, bacteria have a natural tendency to adhere to surfaces as a survival mechanism. Bacterial colonization of solid surfaces has been described as a basic and natural bacterial stratagem in a wide variety of environments (Hunt et al., 2004). Hydrophobicity of the bacterial cell surface is one of the most important factors that govern the mechanism of bacterial adhesion to inanimate biological surfaces (Vesterlund et al., 2005). Although research has been conducted to try to elucidate the molecular and physiological changes associated with Salmonella stress, the mechanisms involved are not completely understood, although it is generally acknowledged that several genes, such as rpoS, and stress proteins (Foster, 2000; Dodd and Aldsworth, 2002) are implied. Probably other macromolecules, such as membrane fatty acids, may also be involved in these bacterial responses. However, few studies, to our knowledge, have been carried out to clarify its potential role. It has been shown that variations in growth temperature (Wang et al., 2005), pH (Wang et al., 2005), ethanol concentration (Teixeira et al., 2002), and external osmolality (Guillot et al., 2000), as well as transition to the stationary phase (Casadei et al., 2002) lead to changes in membrane fatty acid composition. Furthermore, it has been shown that membrane fatty acid composition and membrane fluidity affect the bacterial thermotolerance (Sampathkumar et al., 2004). Annous et al. (1999) showed that Pediococcus spp cells obtained at low temperatures had an increase in the concentration of unsaturated fatty acids of the cytoplasmic membrane, resulting in a higher membrane fluidity, which was linked with a reduced thermal resistance. Sampathkumar et al. (2004) showed an increased thermotolerance in Salmonella Enteritidis when it was pre-treated at alkaline pH values, which are linked to an increased saturated and cyclic to unsaturated fatty acid ratio.
The aim of this work is to study the effects of starvation conditions in seawater microcosm during three years, on the ability of Salmonella Typhimurium cells to adhere to polystyrene microplate, and Hep2 cells. In addition, the membrane fatty acid profile of Salmonella Typhimurium is studied in an attempt to clarify the role of membrane composition and fluidity in Salmonella stress responses.
Methods
Bacterial strains and growth conditions
Salmonella Typhimurium ATCC 14028s (S1) and Salmonella Typhimurium LT2 DT104 (S2), provided from French Food Safety Agency, were used in this study. All strains were maintained at −80°C in Luria-Bertani (LB) broth supplemented with glycerol (15%, vol/vol). For the experiments, the cells were grown at 37°C in tryptic soy broth (TSB; Pronadisa, Spain) for 24 h. The microcosms, natural seawater (100 mL), were filtered through membranes (pore size 0.22 μm; Millipore, Bedford, MA) and autoclaved (121°C/20 min) in 250-mL Erlenmeyer flasks. Salmonella Typhimurium cells were washed three times by centrifugation (13,000 rpm for 10 min at 20°C) with autoclaved seawater and then suspended in 10 mL of autoclaved seawater. The microcosms (100 mL) were inoculated with these suspensions (109 CFU/mL) and then incubated at room temperature for 3 years under static conditions. Three microcosms were used for each strain, and one microcosm without bacteria served as negative control. All the experiments were realized, in triplicate, with stressed and nonstressed bacteria.
Molecular confirmation of stressed bacteria
In order to confirm the starved cells of Salmonella Typhimurium incubated during 3 years in seawater microcosms, polymerase chain reaction (PCR) was used according to the method previously described (Nayak et al., 2004). Bacteria were cultured on trypticase soy agar (TSA) for 24 h at 37°C. One colony was cultured in TSB for 24 h at 37°C, and 1.5 mL was centrifuged. The DNA was extracted by boiling for 5 min and centrifugation at 13,000 rpm for 8 min. The supernatant was used for amplification by PCR with Salmonella primers.
PCR was performed in 25 μL containing 50 ng of extracted DNA, 5 μL green Go Taq buffer (5×), 0.25 μL dNTPs (10 mM), 0.5 μL MgCl2 (50 mM), 1 μL of each InvA forward 5′-TATCGCCACGTTCGGGCAA-3′ and InvA reverse 5′-TCGCACCGTCAAAGGAACC-3′ (25 pM), 1U of GO Taq DNA polymerase (Promega, USA). Amplification was conducted in the Thermocycler PTC 100 (Bio-Rad). The reaction mixtures were heated at 94°C for 5 min and then subjected to 35 cycles of denaturation at 94°C for 1 min, annealing at 57°C for 1 min, and elongation at 72°C for 1 min, followed by 10 min of final extension period at 72°C. PCR products (5 μL) were analyzed on 1% agarose gel stained with ethidium bromide (0.5 mg/mL) at 90 V for 1 h and visualized under ultraviolet transillumination. The amplification products were photographed, and their sizes were determined with a 100-bp molecular size marker (Promega, France).
Adherence assays to polystyrene microplate
The XTT assay was used to quantify bacterial biofilm (Pettit et al., 2005). It measures the reduction of a tetrazolium salt (2,3-bis [2-methyloxy-4-nitro-5-sulfophenyl]-2H-tetrazolium-5-carboxanilide) by metabolically active cells to a colored water-soluble formazan derivative that can be easily quantified colorimetrically (Tunney et al., 2004). XTT (Sigma-Aldrich, Switzerland) solution (1 mg/mL) was prepared in PBS (7 mM Na2HPO4, 3 mM NaH2PO4, and 130 mM NaCl at pH 7.4), filter sterilized and stored at −80°C. Menadione (Sigma-Aldrich, Switzerland) solution (1 mM) was prepared in acetone and sterilized immediately before each assay. An overnight culture grown in TSB (Bio-Rad), of stressed and nonstressed cells, at 37°C was diluted to 1:100 in TSB supplement with 2% (wt/vol) glucose. A total of 200 μL of these cell suspensions was transferred in a U-bottomed 96-well microtiter plate. Each strain was tested in triplicate. Wells with sterile TSB alone served as controls. Following incubation for 24 h at 37°C, the biofilms were first washed five times with 200 μL of PBS, and then 100 μL PBS and 12 μL XTT-menadione solution (12.5:1 v/v) were added to each of the prewashed wells and the control wells. The plate was then incubated for 3 h in the dark at 37°C. Following incubation, 100 μL of the solution was transferred to fresh wells, and the color change in the solution was measured with a multiskan reader (GIO, Rome, Italy) at 492 nm.
Hep 2 cells adherence assays
Quantitative adherence assays of stressed and control Salmonella Typhimurium cells was performed with laryngeal cancer cells (Hep-2) obtained from the American Type Culture Collection (ATCC, Rockville, MD) as described by Chatti et al. (2007). Hep-2 cells were seeded at a concentration of 2×105 cells/mL and grown overnight in minimal essential medium (MEM) with Earle's salts and 10% fetal bovine serum in 96-well microtiter plates at 37°C with 5% CO2. Each Salmonella strain was grown overnight in TSB at 37°C with shaking (150 rpm). The bacterial cells were washed three times by centrifugation at 6000 rpm for 15 min with MEM without serum and resuspended in the same medium. The number of bacteria in the suspension was adjusted to 108 CFU/mL. The monolayers of human cells were inoculated, for each tested strain, with 108 CFU/mL, and incubated at 37°C in 5% CO2 for 60 min. Then, bacterial suspension was removed to exclude the unattached bacteria. The monolayer of Hep-2 cells was washed three times with DMEM, and 1 mL of Triton X-100 in PBS was added for 5 min at room temperature to release the bacteria from the cells. The number of bacteria was estimated by plating serial dilutions. All experiments were performed in triplicate.
Cell surface hydrophobicity
Hydrophobicity was measured by the hexadecane partitioning method of van Loosdrecht et al. (1987). Stressed and control bacterial cells were grown overnight in TSB, and were washed three times by centrifugation at 6000 rpm for 10 min with PBS to remove the broth residues and then resuspended in 4 mL of PBS, and the absorbance (DO540) was determined. One milliliter of hexadecane was added to each cell suspension and vortexed for 5 min. Each suspension was re-incubated at 37°C for 30 min. The aqueous layer was removed and aerated to remove all traces of hexadecane, and absorbance (DO540) was measured against a hexadecane-extracted PBS blank. The hydrophobicity index was expressed as the ratio of absorbance of the hexadecane-extracted sample to absorbance of the sample before extraction.
Analysis of fatty acids
To analyze the total cellular fatty acids, starved and control cells recovered from 10 mL of each cell suspension were pretreated following the MIDI protocols (Sasser, 1990). All reagents for saponification, methylation, extraction, and washing were dispensed with autopipets into this same tube, making the hands-on time minimal. Next, the final extracts were analyzed by gas chromatography (column: 30m×0.25 mm HP-Innowax; flame ionization detect temperature at 280°C; carrier gas N2 at 1 mL/min; injector temperature 270°C; oven temperature programmed from 130°C to 230°C) using a Hewlett-Packard HP 5890 capillary gas chromatograph linked to an HP Chemstation integrator. The identification of fatty acid methyl esters was performed by external standards (all purchased from Sigma Chemical Co.) submitted to the same processes of manipulation as the analyzed biological samples. A known quantity of heneicosanoic acid methyl ester [C21:0] was used as an internal injection standard. The values of fatty acids are presented as area percentage of total fatty acids. Total saturated fatty acids (SFA), total unsaturated fatty acids (UFA), and total cyclic fatty acids (CFA) were used to determine the differences among membrane fatty acids of Salmonella Typhimurium cells grown under the different conditions. The UFA/SFA ratio was used as an indirect indicator of the membrane fluidity.
Statistical analysis
Statistical analysis was performed using the SPSS 13.0 statistics package for Windows. The Friedman test, followed by the Wilcoxon signed ranks test were used to examine differences in the degree of adhesion assay, in the CFA and UFA/SFA. Those p-values of<0.05 were considered as significant.
Results
Molecular confirmation of stressed strains
We used the PCR technique to identify the stressed strains. After amplification of InvA gene by PCR, we confirmed the identity of the investigated Salmonella Typhimurium strains incubated for 3 years in seawater microcosms (Fig. 1).
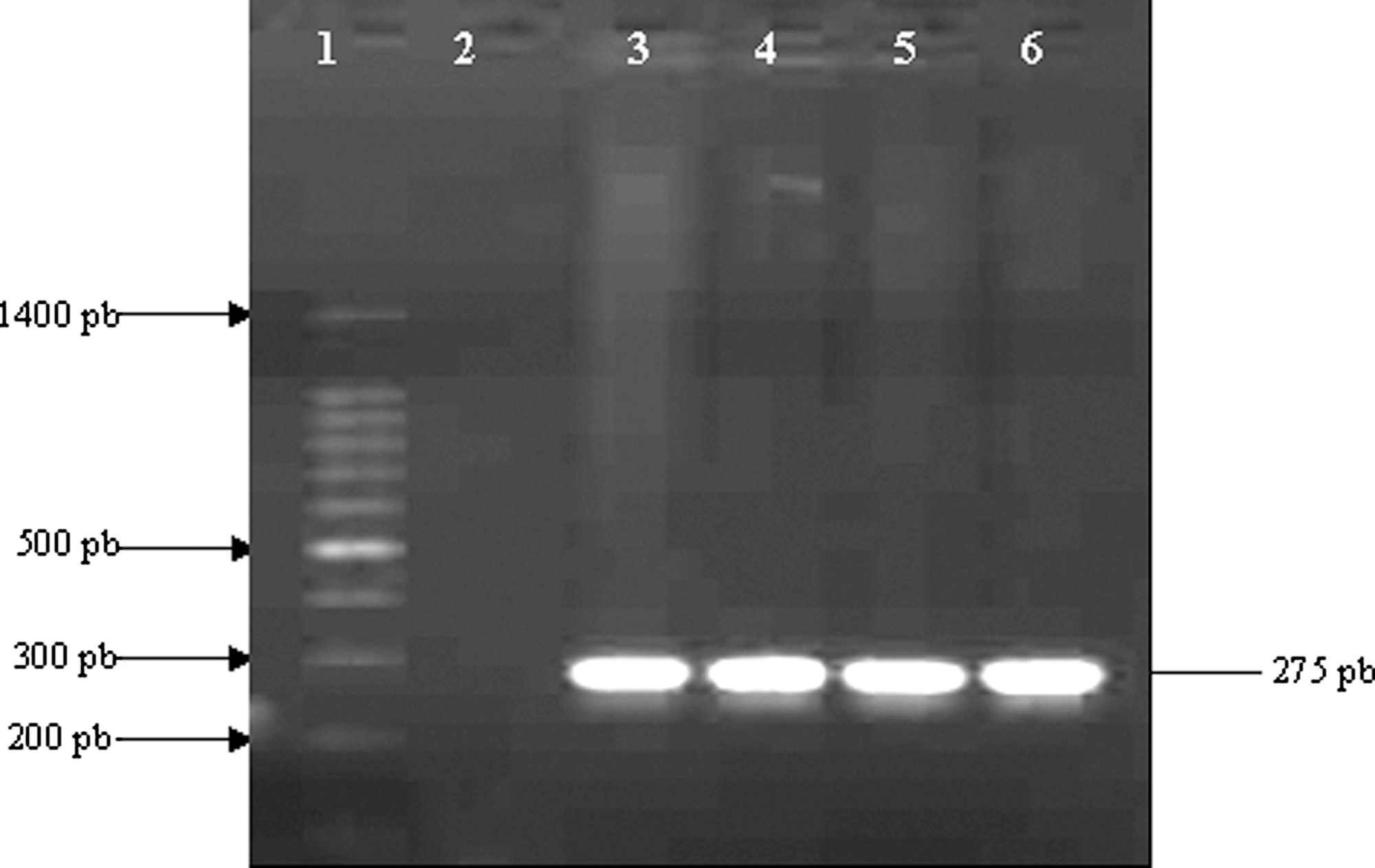
Agarose gel electrophoresis of polymerase chain reaction (PCR) amplification of InvA gene. Lane 1, 100-bp DNA molecular size marker; lane 2, negative control; lanes 3 and 4, S1 and S2, respectively, before incubation; lanes 5 and 6, S1 and S2, respectively, after incubation in seawater microcosm.
Adherence to polystyrene microplate
The two strains of Salmonella Typhimurium were adhesive to polystyrene 96-well microtiter plate at different degrees. The optical density values of XTT reduction estimated at 492 nm were 1.47 for S1 and 1.21 for S2. After incubation in seawater, the ability of starved cells (S1i and S2i) to form biofilm decreased significantly (p<0.05; Table 1).
p<0.05.
S1, Salmonella Typhimurium ATCC 14028s; S2, Salmonella Typhimurium LT2 DT104; I, strain incubated for 3 years in seawater microcosms; SD, standard deviation.
Adherence to Hep-2 cells
Quantitative adherence of Salmonella Typhimurium to Hep-2 cells was assessed before and after incubation in seawater microcosms. All these data are summarized in Table 1. We have noted that S1 and S2 were adherent. Our results showed that, after starvation, the adherence of the two tested strains (S1i and S2i) increased significantly (p<0.05; 4.83% and 6.1%, respectively).
Effect of stress on the surface hydrophobicity
Incubation in seawater had a significant effect on the surface hydrophobicity of Salmonella Typhimurium (Table 1). The cells of the two starved strains (S1i and S2i) showed a significant decrease (p<0.05) in surface hydrophobicity (26.51% and 22.22%, respectively).
Membrane fatty acids analysis
The membrane fatty acid composition of Salmonella Typhimurium before and after starvation was determined using a chromatographic method (Tables 2 and 3). Seventeen fatty acids were found when cells were grown both under normal and stress conditions. The five main peaks were identified as palmitoleic acid (C16:1 w7), palmitic acid (C16:0), oleic acid (C18:1 w9), vaccenic acid (C18:1 w 7), and linoleic acid (C18:2 w 6). Their relative percentages were between 4 and 36, or 78%. Twelve fatty acids were also detected at lower relative concentrations: C10:0, C12:0, C14:0, C14:1, C15:0 iso, C15:0 antieso, Cyc 17, C17:1, C18:0, C18:1 w 7, C20:1 w 9, and C20:0. As expected, incubation of Salmonella Typhimurium in seawater resulted in differences in membrane fatty acid composition. We observed a significant increase in the proportion of CFA for S1i (p<0.05) and a significant decrease for S2i compared to controls (p<0.05). As shown in Table 3, the UFA/SFA ratios observed for stressed cells (S1i, S2i) were significantly lower (1.45; 1.40) than those of control cells (S1, S2; p<0.05).
S1, Salmonella Typhimurium ATCC 14028s; S2, Salmonella Typhimurium LT2 DT104; i, strain incubated for 3 years in seawater microcosms; C10:0, capric acid; C12:0, lauric acid; C14:0, myristic acid; C14:1, myristoleic acid; C15:0 iso, isopentadecylic acid; C15:0 antieso, antiesopentacyclic acid; C16:0, palmitic acid; C16:1w7, palmitoleic acid; Cyc 17, methylenhexadecanoic acid; C17:1, heptadecenoic acid; C18:0, stearic acid; C18:1w9, oleic acid; C18:1w7, vaccenic acid; C18:2w6, linoleic acid; C18:3w6, gamma-linolenic acid; C20:0, arachidic acid; C20:1w9, eicosenoic acid.
p<0.05.
S1, Salmonella Typhimurium ATCC 14028s; S2, Salmonella Typhimurium LT2 DT104; i, strain incubated for 3 years in seawater microcosms; SFA, total saturated fatty acids; UFA, total unsaturated fatty acids; CFA, total cyclic fatty acids; UFA/SFA, unsaturated to saturated ratio.
Discussion
Bacterial cells have the ability to sense and respond to changes in their external environment (Rosen and Belkin, 2001). This ability of bacteria is crucial for their survival (Ben Abdallah et al., 2009). Salmonella Typhimurium encounters many diverse and extreme environments, and as a result it has developed responses to combat these adverse conditions (Foster and Spector, 1995). The results developed in the present work show that Salmonella Typhimurium are able to adapt and survive under extremely stressful conditions. The persistence of Salmonella under starvation and/or osmotic stress induced modifications in cell surface properties. Our study showed that the capacity of Salmonella Typhimurium cells to adhere to polystyrene and their hydrophobicity were influenced by nutrient deficiency and/or osmotic stress. Indeed, the starved cells in seawater formed significantly less biofilm on polystyrene thus correlated with a significant decrease on their hydrophobicity. These results are in accordance with those reported by Ngwai et al. (2006), who found a significant reduction in biofilm amount formed on polystyrene when both Salmonella Typhimurium and Salmonella Enteritidis were cultured in the starvation medium. Starvation is known to alter bacterial surface characteristics such as hydrophobicity, charge, and irreversible attachment, which are essential factors in biofilm formation (Brown et al., 1977; Kjelleberg and Hermansson, 1984). Various studies suggest that fimbrial expression is associated with biofilm formation. Indeed, fimbriae are known to be involved in attachment (Edwards and Schifferli, 1997) and biofilm development (O'Toole and Kolter, 1998). Furthermore, previous studies have shown that regulation of fimbrial production is affected by carbon and nitrogen sources (Edwards and Schifferli, 1997), pH (Low et al., 1996), and temperature (Xie et al., 1997). Thus, we suggest in the present case, a modulation of the fimbrial expression affecting the biofilm formation in seawater microcosm. Our results showed that the ability of Salmonella Typhimurium to adhere to Hep-2 cells has increased under starvation and osmotic stress. This finding correlates with other reports on Shigella strains, showing that its adhesive power on KB cells increased after incubation in seawater (Ellafi et al., 2009). In the same way, Tartera and Metcalf (1993) reported that the adhesion capacity of Salmonella to human intestinal cells increased in relation to the increase of the medium concentration with NaCl. Other studies have shown that exposure to stress, including acid shock, enhance adhesion of H. pylori, L. pneumophila, and C. difficile (Huesca et al., 1996; Waligora et al., 1999).
Based on our results, it appears that starvation is the main cause of changes in membrane fatty acids composition of Salmonella Typhimurium. The decrease of the UFA/SFA ratio mainly due to the decrease of C18:1 may contribute to the membrane adaptation of Salmonella Typhimurium to starvation. Previously, similar results have proven that low UFA/SFA ratio was linked to less membrane fluidity (Casadei et al., 2002; Wang et al., 2005). It is generally admitted that cells regulate their lipid composition in order to achieve a degree of fluidity compatible with life (Teixeira et al., 2002). One of the most important consequences of membrane fatty acid changes in microorganisms is to modulate the activity of intrinsic proteins that perform functions such as ion pumping and nutrient uptake (Russell and Fukanaga, 1990). Furthermore, it has been demonstrated that alterations of the lipid composition in Pseudomonas aeruginosa occured during the transition period from the late exponential phase to the stationary phase. This seems to be a strategy used by the cells to conserve energy by reducing membrane fluidity (Costerton et al., 1999). In addition, Moorman et al. (2008) have shown that cell surface properties of Listeria innocua, including cell hydrophobicity and membrane fluidity as impacted by fatty acid composition, can be altered through exposure to various environmental stresses including cold, heat acid, and starvation. Further, membranes fatty acids can change in length, saturation, and branching in response to stress, and these changes alter membrane fluidity and the cell's ability to interact with and survive in the surrounding environment (Voet and Voet, 1995). In this work, we also showed that cyc 17 CFA production was significantly affected by stress conditions. This modification was manifested by an increase in the Salmonella Typhimurium ATCC 14028s and a decrease in Salmonella Typhimurium LT2 DT104. However, CFA contribution to membrane properties is not yet understood, especially concerning the modifications of membrane fluidity in response to environmental stress. According to Duforc et al. (1984), the presence of a cyclopropane ring within membrane fatty acid increases the stability of the structural and dynamic properties of biological membranes. Other studies have suggested that an increase in CFA content could cause a decrease in UFA/SFA ratio and in membrane fluidity (Magnuson et al., 1993; Annous et al., 1999), which could explain the increase in thermal resistance of Salmonella Typhimurium cells with high CFA levels. In addition, Brown et al. (1997) have shown that CFA play an important role in acid adaptation responses in E. coli.
In conclusion, starvation and/or osmotic stress influence the capacity of biofilm production and the surface hydrophobicity of Salmonella Typhimurium. Despite these changes in surface proprieties, adhesion into Hep-2 cells is enhanced in starved cells. In addition, membrane fatty acids composition was also modified. This can play an important role in the survival of this germ under these stress conditions. These findings lead us to hypothesize that stress enhance virulence by increasing host cell adhesion. These results are very important in terms of explaining the pathogenesis of this bacterium.
Footnotes
Disclosure Statement
No competing financial interests exist.