
Abstract
Plasmid-mediated quinolone resistance (PMQR) determinants were widely distributed among Enterobacteriaceae. The objectives of the present study were to analyze PMQR-positive Escherichia coli isolates from pigs, and to investigate the association between these determinants and other resistant genes. A total of 129 porcine E. coli isolates were included in this study. The presence of PMQR, floR, bla CTX-M-14, and bla TEM-1 genes were detected by polymerase chain reaction (PCR) amplification and confirmed by subsequent sequencing. The PMQR-positive isolates were subjected to plasmid profiling, and transformation experiments were conducted to identify the quinolone resistance plasmids. The qnrS1 region of a quinolone resistance plasmid was cloned and sequenced. Among the 129 E. coli isolates, the positive rate for PMQR determinants was 42.6%, and the prevalence of qnr genes, aa(6')-Ib–cr, and qepA were 23.3%, 18.6%, and 0.8%, respectively. A qnrS1-carrying plasmid of 81 kb, named plasmid T078 (pT078), was detected from one multidrug-resistant isolate. Hybridization and PCR analysis confirmed that floR, bla CTX-M-14, and bla TEM-1 genes were also located on this plasmid. Sequence analysis identified the qnrS1 gene flanked by a truncated transposase gene. Moreover, complete tetracycline resistance genes tet(A) and tet(R) were found upstream of the qnrS1 gene, and floR gene was found downstream of the qnrS1 gene on the plasmid pT078. To our knowledge, this is the first study demonstrating the occurrence of qnrS1, floR, bla CTX-M-14, bla TEM-1, and tet(A) on one plasmid in E. coli isolated from food animals.
Introduction
S
The ability of plasmids to evolve independently of their hosts has allowed numerous resistance genes from diverse species of bacteria to be assembled within a single plasmid and transformed into a wide variety of organisms (Galani et al., 2010). There have been many reports on the association of the qnrA determinants with ESBLs (VEB-1, SHV-7, and CTX-M) (Ferreira et al., 2010; Nordmann and Poirel, 2005). The qnrB gene was also found to co-occur with the carbapenemase gene bla KPC-2 on the same plasmid (Chmelnitsky et al., 2008; Endimiani et al., 2008). However, other resistance genes were seldom reported to co-occur with the qnr genes. Our previous study showed that the bla CTX-M genes, floR and qnr genes, were highly prevalent in E. coli isolated from animals in China (Huang et al., 2011; Xia et al., 2011).
The objectives of the present study were to examine the emergence of potential plasmid-mediated quinolone resistance (PMQR) determinants coexistent with bla CTX-M genes and floR gene in E. coli isolates from pigs in China and to identify the association between qnr genes and other resistance genes.
Methods
Bacterial isolates and susceptibility testing
Bacterial strains were obtained from three pig farms in North China by the China Institute of Veterinary Drug Control and five pig farms in South China by Sichuan Agricultural University from 2005 to 2007. Samples were collected from feces of pigs with diarrhea using sterile cloacal swabs. The samples were inoculated in sterile nutrient broth (Becton Dickinson Difco Laboratories, Detroit, MI) and incubated for 12 h at 37°C. The strains from the same pig were grown on one chromogenic medium plate for selecting E. coli (CHROMagar, Paris, France). Only one colony per plate was selected, and blue colonies on the culture plates were regarded as presumptive E. coli colonies. A total of 129 E. coli isolates were confirmed using the API-20E system (bioMerieux, Marcy l'Etoile, France). Minimum inhibitory concentrations (MICs) of isolates and transconjugants were performed by the broth microdilution method in accordance with Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2007, 2008). E. coli ATCC 25922 was used as a quality control strain (Lu et al., 2011).
PCR amplification and DNA sequencing
Rapid DNA preparation was performed by heating a single colony at 100°C in a volume of 100 μL of distilled water, followed by a centrifugation step to remove cell debris. Based on sequence alignments of the qnrA-, qnrB-, qnrS-, qnrC-like genes; qepA; and aac(6')-Ib, polymerase chain reaction (PCR) primers were designed to amplify internal fragments of the target genes with sizes of 580, 264, 428, 307, 218, and 482 bp, respectively (Table 1). PCR conditions were as follows: (i) 5 min at 95°C and (ii) 35 cycles, with 1 cycle consisting of 30 s at 94°C, 40 s at 56°C, 1 min at 72°C, and (iii) 10 min at 72°C. For the PMQR-positive isolates, amplification and identification of ESBL-encoding genes (bla TEM , bla CTX-M) and floR gene were performed with previously described primers (Table 1) (Blickwede and Schwarz, 2004; Huang et al., 2009). PCR experiments were carried out using the above conditions with an annealing temperature of 56°C, 58°C, or 60°C, respectively. Both strands of amplicons were sequenced using an ABI 3730 sequencer (Applied Biosystems, Norwalk, CT) with the same primers used for PCR amplification.
Transfer of PMQR determinants
Conjugation experiments using the azide-resistant E. coli strain J53AzR as the recipient were performed as described previously (Wang et al., 2003). Transconjugants were selected on trypticase soy agar (TSA) plates containing sodium azide (100 mg/L) and ampicillin (100 mg/L). Transformation of E. coli DH10B with the plasmid DNA of clinical E. coli isolates was performed using a standard electroporation technique. Transformants were selected on TSA plates containing ciprofloxacin (0.04 mg/L).
Southern hybridization analysis
The QIAGEN Large-Construct Kit (Qiagen, Hilden, Germany) was used to extract plasmids from floR-positive isolates, bla CTX-M-positive isolates, and transformants according to the manufacturer's instructions. E. coli V517, E. coli J53 R27, E. coli J53 R1, and E. coli J53 Plac were used as plasmid size standards. Southern hybridization using DIG DNA labeling and detection kit (Roche Applied Science, Mannheim, Germany) was performed with PCR-generated probes specific for qnrS1, bla TEM-1, bla CTX-M-14, or floR probes.
Cloning and sequence analysis of the qnrS1 region of plasmid pT078
Plasmid pT078 extracted from transformant T078 was digested with EcoRI, ligated to the same restricted pBluescript II SK+ (Stratagence, Amsterdam, The Netherlands), and transformed into E. coli DH10B. Selection of clones carrying the qnrS1-containing fragment was performed on Luria agar plates containing ampicillin and nalidixic acid. Restriction enzyme analysis was used to determine the size of the cloned qnrS1-carrying fragments. The nucleotide sequences of the qnrS1-bearing fragments were first determined by using the commercially available standard primers M13 universal and M13 reverse. Additional sequences flanking the qnrS1-carrying region were then determined by primer walking using the non-digested plasmid as a sequencing template. The sequence of a 17575-bp segment of plasmid pT078 has been deposited in the GenBank database under the accession number GQ214053. Sequence comparisons were carried out using the BLAST program, available at the U.S. National Center for Biotechnology Information (
Results
Screening and identification of resistance genes
Among the 129 E. coli isolates examined in this study, the prevalence of qnr genes, aac(6')-Ib–cr, and qepA were 23.3% (30), 18.6% (24), and 0.8% (1), respectively. Among the qnr determinants, qnrA-, qnrB-, and qnrS-type genes were detected in one (0.8%), seven (5.4%), and 22 (17.1%) strains, respectively, but no isolate was positive for qnrC. Among the 55 PMQR-positive E. coli isolates, the positive rate for bla TEM-1 genes was 100%. Strain DS05, 1206-S23, and 1205-S13 contained both qnrS gene and floR gene. Strain 1213-S18 and 1207-SE51 contained both qnrS gene and bla CTX-M-15 gene. Interestingly, strain 1205-S13 also contained bla CTX-M-14 gene (Table 2).
The strains starting with “T” are transformants.
NAL, nalidixic acid; CIP, ciprofloxacin; ENR, enrofloxacin; NOR, norfloxacin; OFL, ofloxacin; CAZ, ceftazidime; CTF, ceftiofur; GEN, gentamicin; AMK, amikacin; FFC, florfenicol; DOX, doxycycline; AMP, ampicillin; NA, not applicable; MP, multiple plasmids.
Transferability of resistance determinants
For the PMQR-positive isolates, such as strains DS05, 1206-S23, 1205-S13, 1213-S18, and 1207-SE51, conjugation experiments were performed with E. coli J53 as the recipient strain by liquid and solid mating-out assays as described previously (Wang et al., 2003). Transformation of E. coli DH10B with the plasmid DNA extracted from clinical E. coli isolates was performed using a standard electroporation technique, and the five transformants were named T045, T075, T078, T065, and T073, respectively. Results of antimicrobial susceptibility testing for the five donors and five transformants were shown in Table 2, and the antimicrobial susceptibility of the five transconjugants was similar to the five transformants (data not shown). Molecular analysis revealed that all of the five donors had multiple plasmids, and each of the transformants carried only one transferable plasmid (Table 2). The floR gene in 1205-S13 was cotransferred to T078 with qnrS1 gene, whereas the floR genes in DS05 and 1206-S23 were not. The bla CTX-M-14 gene in 1205-S13 was also cotransferred to T078 with qnrS1 gene, whereas the bla CTX-M-15 genes in 1213-S18 and 1207-SE51 were not. Electrophoresis of the plasmid (pT078) extracted from transformant T078 was followed by Southern hybridization as described previously. The result showed that the four genes (qnrS1, bla TEM-1, bla CTX-M-14, and floR) were located on the same plasmid (see Fig. 2 below). The ciprofloxacin MICs of the transconjugants and transformants were 0.5 mg/L, representing a 64-fold increase over that of the recipient strain (MIC 0.008 mg/L). Increases in the MICs of multiple classes of antimicrobials were observed in the transformants, including a 32-fold increase in the MICs of three FQs from 0.25 to 0.5 mg/L (Table 2).
Analysis of the qnrS1-carrying plasmid pT078
The 81.2-kb plasmid pT078 mediated resistance to florfenicol, ciprofloxacin, and ceftiofur as well as to doxycycline, which was confirmed by in vitro susceptibility testing of transformants. PCR for the qnrS, floR, bla CTX-M-14, and bla TEM-1 genes showed that all of these genes were present on the pT078 plasmid. A 17575-bp segment of pT078, which comprised the tet(A), qnrS1, and floR regions, was cloned and sequenced. The open reading frames (ORFs) detected in this segment are displayed in Figure 1. The sequence consisted of three different segments; the initial 5515-bp exhibited 99% nucleotide sequence identity to the plasmid pWSH1 (accession no. DQ464880). Within this region, tet(A) resistance gene, tet(R) gene, pecM gene, and an incomplete tnpA gene were detected. The adjacent 4614 bp showed 100% identity to plasmid pINF5 from Salmonella Infantis (Kehrenberg et al., 2006a) and included the qnrS1 resistance gene area, two truncated CS12 insertion sequences, and an incomplete Tn5058-related resolvase gene. Further downstream, another 7096-bp region with 98% identity to plasmid pAR060302 (accession no. FJ621588) was detected. This region included floR resistance gene, a truncated IS26 insertion sequence, and three ORFs. Between the last two segments, a 350-bp region without significant homology to database entries was detected (Fig. 1).
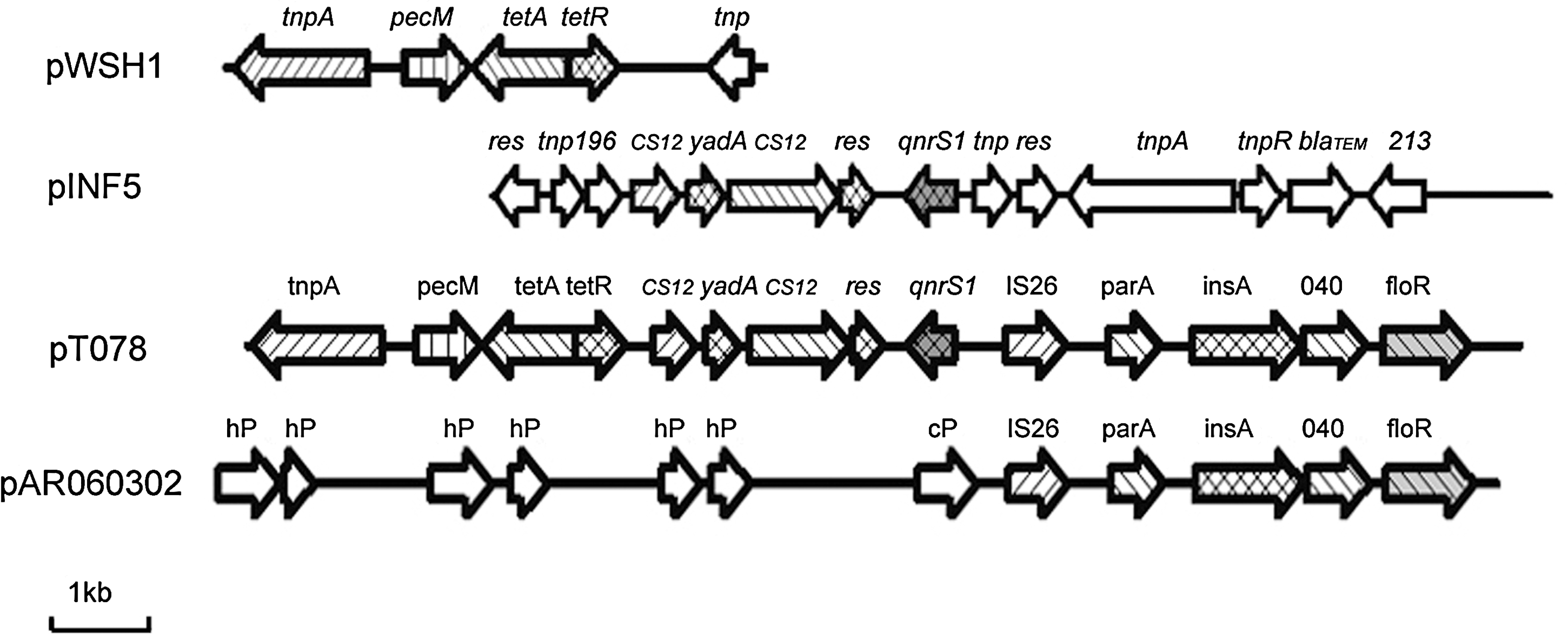
Comparison of the sequenced parts of plasmids pWSH1, pINF5, and pAR060302 with the fragment of pT078 from Escherichia coli. The reading frames are shown as arrows, with the arrowhead indicating the direction of transcription. The arrows marked as hP and cP in the map of pAR060302 indicate reading frames for hypothetical proteins and a conserved protein, respectively. The areas of homology are shown by using the same shape.
Restriction analysis of pT078 was performed using three endonucleases, respectively. Restriction fragments generated with EcoRI, XbaI, or BamHI were chosen for Southern hybridization with qnrS1, bla TEM-1, bla CTX-M-14, or floR probes. In the case of restriction with EcoRI, qnrS1, and floR probes hybridized to the same fragment (18kb) (Fig. 2), for the lines which were digested by XbaI, all of the probes hybridized to the same fragment (23 kb), and the undigested plasmid (Fig. 2).

Restriction and hybridization analysis of the plasmid pT078.
Discussion
The use of antibiotics in the prophylaxis of diseases as well as their effect on growth promotion of healthy animals is common in livestock production in China. The emergence of antibiotic resistance of pathogens is a growing concern in veterinary medicine. Antibiotic-resistant pathogens pose the threat of severe and costly animal health problems. Furthermore, the increasing level of resistance to frontline antimicrobial agents in treating human diseases, such as extended-spectrum cephalosporins and fluoroquinolones, is a significant public health concern (Krueger et al., 2011).
The widespread dissemination of PMQR could potentially fuel the rapid development of fluoroquinolone resistance, because PMQR reduces the susceptibility to antibiotics and supports the occurrence of resistance-associated mutations on bacterial chromosomes. To date, these PMQR genes have been widely reported in Southern and Eastern Asia, North and South America, and Europe (Amabile-Cuevas et al., 2010; Dahmen et al., 2010). These PMQR determinants are usually carried on mobile elements and can be transferred among different bacterial strains or species. In isolates from Canada, France, Thailand, and Turkey, qnrA1 was associated with VEB-1β-lactamases (Poirel et al., 2005), while in samples from other countries, qnrA1 and ESBL SHV-12 and various CTX-M enzymes were linked (Bado et al., 2010). The qnrB alleles had also been associated with, or linked to the ESBLs SHV-12, TEM-52, and various CTX-M (Cattoir et al., 2007; Jiang et al., 2008). Unlike qnrA and qnrB, qnrS was seldom associated with ESBL or AmpC β-lactamase genes. In the present study, we identified bla CTX-M-14, bla TEM-1, floR, and qnrS1 genes located on one plasmid. Then, we characterized a large fragment carrying the qnrS1 gene and floR gene. Unexpectedly, another resistance gene, tet(A), was also located on this fragment. To our knowledge, this is the first study showing the occurrence of qnrS1, floR, bla CTX-M-14, bla TEM-1, and tet(A) located on the same plasmid isolated from food animals. In addition to the potential of the strain to develop high-level resistance to quinolones, the coexistence of multiple resistance genes on the same plasmid, which may confer resistance to β-lactamases, tetracycline, florfenicol, and quinolones, poses a serious epidemiological, clinical, and public health threat. The horizontal dissemination of plasmids carrying multiple antimicrobial resistance genes is rapidly resulting in more bacterial strains that are resistant to commonly used antibiotics. Antibiotic resistance developed in food-producing animals can be transferred to humans through food consumption or direct contact, and this has been shown in several studies (Manian, 2003; Molbak et al., 1999). The continuous and widespread use of florfenicol, fluoroquinolones, and β-lactamases in animal production will maintain the long-term presence of these resistance gene linkages. Since E. coli has a broad host range, a more large-scale appearance of these resistance gene linkages among different strains in both humans and animals is predicted for the future.
Footnotes
Acknowledgments
We would like to thank George A. Jacoby, David C. Hooper, and Minggui Wang for kindly providing the E. coli strains J53AzR, V517, R1, R27, and plac. We thank Dr. Thomas Vorup-Jensen for assisting with the writing of this manuscript. This study was supported by the Program for Cheung Kong Scholars and the Innovative Research Team at the University of China (grant IRT0866), and Exclusive Research Fund for Public Welfare from Ministry of Agriculture of the People's of China (grant 200903055).
Disclosure Statement
No competing financial interests exist.