
Abstract
Strains belonging to Bacillus cereus Group include six different species, among which are Bacillus thuringiensis, Bacillus weihenstephanensis, and Bacillus cereus sensu stricto, a causative agent of food poisoning. Sequence of the panC-housekeeping gene is used for B. cereus Group affiliation to seven major phylogenetic groups (I–VII) with different ecological niches and variations in thermal growth range and spore heat resistance of B. cereus Group microorganisms varies among phylogenetic groups. We assigned a selection of B. cereus sensu stricto strains related to food poisoning from the Spanish cultivar Collection (Valencia) to Group IV strains based on panC gene sequence. Thermal inactivation assays revealed variability of spore heat resistance within these Group IV strains. Adequate food sanitizing treatments therefore require fast and reliable identification of particular strains. In the present study, feasibility of genotyping via high-resolution melting (HRM) analysis was examined. HRM analysis of amplified polymorphic 16S-23 intergenic spacer region (ISR) region proved to be discriminatory for B. cereus sensu stricto strain typing, while two other polymorphic regions within the bacterial rRNA operon allowed differentiation between Bacillus species, demonstrating its applicability for discrimination on the species and strain level within B. cereus Group.
Introduction
T
Guinebretière et al. (2008) reported on an “ecotypic” structure of populations in the B. cereus Group with a differentiation into seven phylogenetic groups based on phenotypic and genetic criteria. Every group shows a specific “thermotype” and virulence potential with implications for the food industry. B. cereus sensu stricto strains are affiliated to groups II, III, IV, and V, making differentiation at the strain level an important issue, since toxins produced by Bacillus species were reported as causative agent in 124 outbreaks affecting 1,132 people and causing 93 hospitalizations during 2009 (EFSA, 2011).
All members of the B. cereus Group show a very high similarity level (from 99.5% to 100%) of the 16S and 23S rRNA sequences (Daffonchio et al., 1998) and a variable intergenic spacer region (ISR) due to nucleotide substitutions, insertions, deletions, and presence of tRNA genes (Cherif et al., 2003). The 16S-23S ISR usually shows significant polymorphisms at genus and species level, and intra-species variations on rRNA operon genes can be detected in some cases by conventional polymerase chain reaction (PCR) in combination with agarose gel electophoresis (Daffonchio et al., 1998).
Differentiation of closely related species and strains of the B. cereus Group remains a challenge which could be addressed using high-resolution melting (HRM) analysis, an extension of previous DNA dissociation (or “melting”) analyses based on highly controlled temperature transitions and DNA binding dyes with improved saturation properties, allowing the detection of single base sequence variations. HRM has been reported to obtain successful results in specific species identification of clinically relevant bacterial species (Yang et al., 2009) and in medical diagnosis applications (Reed et al., 2007; Montgomery et al., 2007; Erali et al., 2008; Van Ert et al., 2007).
The aim of this investigation was to assign thermal niches to five selected B. cereus sensu stricto strains from the Spanish Type Culture Collection (CECT), based on partial panC gene sequencing and heat resistance. They were classified by Martínez-Blanch et al. (2009) into three groups with different virulence factors. Our results showed that the bacterial isolates analyzed belonged to B. cereus Group IV but with variations in heat resistance. We therefore investigated the feasibility of HRM analysis to discriminate B. cereus at the strain and species level. As proof of concept, we analyzed B. cereus CECT131, CECT148, CECT193, CECT4014, and CECT4094, B. weihenstephanensis, and B. thuringiensis strains amplifying a 16S-23S ISR region as well as two other polymorphic areas within the rRNA gene operon.
Our study is a combined approach that includes strain and species identification with phylogenetic affiliation of isolates and characterization of their spore heat resistance.
Methods
Bacterial strains and growth conditions
B. cereus sensu stricto strains used in this study were selected from the Spanish Type Culture Collection (CECT, Valencia, Spain). B. weihenstephanensis (KBAB4) and B. thuringiensis (Bt407) strains were provided by INRA (Avignon, France) (Table 1).
CECT, Spanish Type Culture Collection; INRA, National Institute of Agronomy Research, France.
Following CECT indications, CECT131, CECT148, CECT193, and CECT4094 strains were grown at 30°C and CECT4014 strain at 37°C in nutrient broth for 18 h and stored with 20% (v/v) glycerol at −80°C. For DNA extraction, depending on the strains, cultures were grown in brain-heart infusion (BHI) broth at 30°C and 37°C, as described above, for 18 h with agitation at 200 rpm. Strains were sporulated in Fortified Nutrient agar at 30°C (Mazas et al., 1995) and stored at −20°C until use. Spores were heated at 80°C for 10 min prior to use (Fernández et al., 2001).
DNA extraction
Genomic DNA for panC gene sequence analysis was extracted from pure cultures of each B. cereus strain. Cells were lysed in 400 μL of extraction buffer (2% Triton X-100, 1% sodium dodecyl sulfate [SDS], 100mM NaCl, 10mM Tris pH 8, and 1mM ethylenediaminetetraacetic acid [EDTA] pH 8.5, 0.5 mg μL−1 lysozyme, 0.023 mg μL−1 proteinase K) (Manzano et al., 2003; Candelon et al., 2004), during 30 min at 37°C, and after adding SDS (1% final concentration), for further 30 min at 65°C. DNA purification included phenol-chloroform-isoamyl alcohol (25:24:1) extraction, followed by isopropanol precipitation; then DNA was finally resuspended in 40 μL of sterile distilled water. Genomic DNA for real time–polymerase chain reaction (RT-PCR) and HRM-PCR was isolated using the DNeasy Blood and Tissue kit (Qiagen, Germantown, MD).
PanC amplification and sequencing
Primers for the amplification of panC gene of the B. cereus Group species were designed with Primer3 (version 0.4.0) software available at
Phylogenetic analysis and affiliation to B. cereus Group
Nucleotide sequence alignement and phylogenetic analysis of panC gene between microorganisms of the B. cereus Group was performed with the ClustalX 2.1 program applying the Neighbor-Joining method (Saitou and Nei, 1987). The analyzed data set also included panC gene sequences of Bacillus species and strains available from GenBank (Benson et al., 2004).
Applying the classification system proposed by Guinebretière et al. (2008), panC gene sequences were used to assign the strains to phylogenetic groups I–VII performing the homology search algorithm offered at
Heat resistance determination of B. cereus sensu stricto spores
Heat resistance determination of B. cereus sporulated cells at 90°C, 95°C, and 100°C on sterile distilled water was carried out using a Mastia thermoresistometer (Conesa et al., 2009). Samples were collected into sterile tubes, diluted, plated onto BHI agar, and incubated at 37°C for 24 h. Survival curves were fitted using GInaFIT (Geeraerd et al., 2005). Maximum inactivation rate and specific inactivation rate (1/min), both designed as Kmax, and shoulder length (min) were obtained when appropriate. Root mean sum of squared errors (RMSSE) (Ratkowsky, 2003) was selected as a measure of goodness-of-fit.
Identification of highly polymorphic genomic regions and primer design
Genome sequences of B. cereus strains with the following accession numbers were obtained from GenBank database and used for primer design: NC 004722 B. cereus ATCC 14579 and NC 0011772 B. cereus G98421. Both sequences were aligned using the multiple sequence alignment algorithm MUSCLE, version 3.8 (
Amplification of polymorphic region by conventional and real-time PCR
The highly polymorphic 16S-23S ISR regions were amplified by conventional PCR using primers 16S-23SISRfor and 16S-23SISRrev (Table 2) from CECT 131, CECT 148, CECT 193, CECT 4014, CECT 4094, Bt407, and KBAB4. The PCR was performed with Herculase®II Fusion Enzyme kit according to manufacturer's instructions. PCR amplicons were analyzed on 2% agarose gels.
Real-time PCR for the region 16S-23S ISR and for polymorphic regions flanking 5S and 16S rRNA genes was performed for the B. cereus Group strains described above, as well as for Bt407 and KBAB4. The 25-μL PCR reaction contained 12.5 μL of Brilliant II SYBR Green QPCR master mix (Stratagene), 0.3μM of forward and reverse primers 16S-23SISR, 5S-GT, and 16S-ISR (Table 2) and template DNA (50–150 ng). Amplifications were performed in triplicate using Rotor-gene Q thermal cycler (Qiagen). PCR conditions consisted of one cycle at 95°C for 10 min, followed by 40 cycles at 95°C for 30 s, 57°C (ISR) or 58°C (5S-GT and 16S-ISR) for 30s and 72°C for 1 min. Melting curve analysis was performed from 57°C to 95°C increasing by 0.1°C each step. Data were analyzed using the software Rotor-Gene 2.02.4.
HRM of polymorphic regions
For the detection of sequence polymorphisms, we performed HRM of 16S-23S ISR region and of polymorphic regions flanking 5S and 16S rRNA genes from CECT 131, CECT 148, CECT 193, CECT4014, CECT 4094, Bt407, and KBAB4 using Type-it® HRM kit (Qiagen) according to the manufacturer's instructions. PCR runs prior to HRM analysis were performed in triplicate using Rotor-gene Q thermal cycler and repeated three times. PCR conditions consisted in one cycle at 95°C for 10 min, followed by 40 cycles at 95°C for 30 s, 57°C (ISR) or 58°C (5S-GT and 16S-ISR) for 30s, and 72°C for 1 min. HRM curve analysis was performed in a temperature range from 57°C (ISR) or 58°C (5S-GT and 16S-ISR) to 95°C increasing 0.1°C each step. Rotor-Gene 2.02.4 software was used to run HRM-PCRs and analyze results. Raw data of HRM analysis were normalized by setting the same start and ending fluorescent signal level for all the curves through adjusting two normalization regions, one region for the start (trailing range) and the other for the end (leading range). Threshold values for genotyping confidence percentages were set. Genotypes were determined automatically by the software, establishing confidence values as an integrity check of obtained results. Samples below 90% threshold value were considered as a genotypic variant.
Results
PanC gene sequence affiliation and phylogenetic analysis of panC gene
Affiliation of B. cereus strains CECT131, CECT148, CECT193, CECT4014, and CECT4094 to the phylogenetic B. cereus Groups I–VII was performed by amplification and sequencing of the panC gene (Candelon et al., 2004; Guinebretière et al., 2008). Results indicated that all tested strains belong to group IV with a 100% homology score. Phylogenetic analysis with tree construction (Fig. 1) for B. cereus CECT strains with those available at the NCBI GenBank is consistent with results of Guinebretière et al. (2008). Results revealed that CECT B. cereus strains are closely related to other strains of the same species as well as B. thuringiensis and B. weihenstephanensis strains.
Genetic relationship among Bacillus cereus Group strains based on panC gene. Neighbor-Joining method (Saitou and Nei, 1987) was used for inferring the topology. Bootstrap values (10000 bootstrap replicates) are shown in the major nodes. Strains' affiliation into major phylogenetic groups is designated by VII, VI, IV, and III, which correspond to phylogenetic groups VII, VI, IV, and III, respectively.
Heat inactivation of B. cereus strains affiliated to group IV
We characterized the heat resistance of strains CECT 131, CECT 148, CECT 193, CECT 4014, and CECT 4094 under isothermal conditions. Results include shoulder length, K max values, and RMSSE for non-linear models (Table 3). Strain CECT 4094 was the most heat resistant, followed by CECT 131 and CECT 148, whereas strains CECT 193 and CECT 4014 are the most heat-sensitive ones (Fig. 2A).
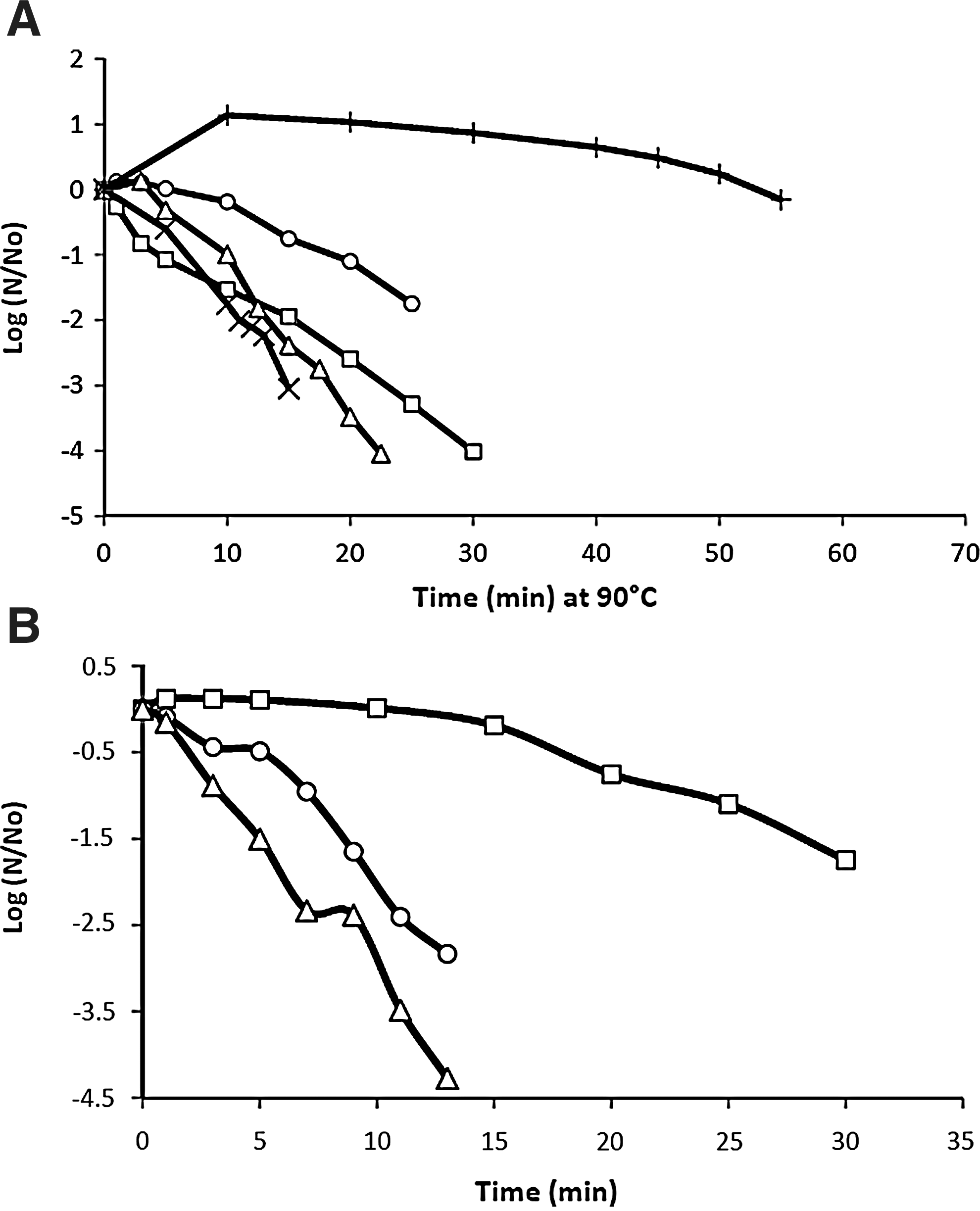
Survival curves at 90°C of Bacillus cereus Spanish Type Culture Collection (CECT) 131 (•), B. cereus CECT 148 (▪), B. cereus CECT 193 (▴), B. cereus CECT 4014 (x) and B. cereus CECT 4094 (+)
Sl, shoulder length; K max, maximum inactivation rate or specific inactivation rate; RMSSE, root mean sum of squared errors.
Shoulder phenomena lower than 0.5 min were considered not significant. At 90°C, strain CECT 4094 presented a shoulder phenomenon with values of 23.88–31.19 min, strain CECT 131 of 9.35–10.93 min, and strain CECT 4014 of 2.68–3.51 min. Shoulder length diminished at higher temperatures in a range of 2.4–4 times for strain CECT 4094 at 95°C and 100°C, whereas for strains CECT 131 and CECT 4014, shoulder phenomenon was not significant (Fig. 2B). Shoulder phenomena are widely described (van Zuijlen et al., 2010).
16S-23S ISR PCR and Tm profiles for strain and species discrimination
Amplification of 16S-23S ISR in B. cereus CECT131, CECT148, CECT193, CECT4014, CECT4094, B. weihenstephanensis KBAB4, and B. thuringiensis Bt407 by conventional PCR resulted in identical and reproducible patterns in all strains tested (Fig. 3A).
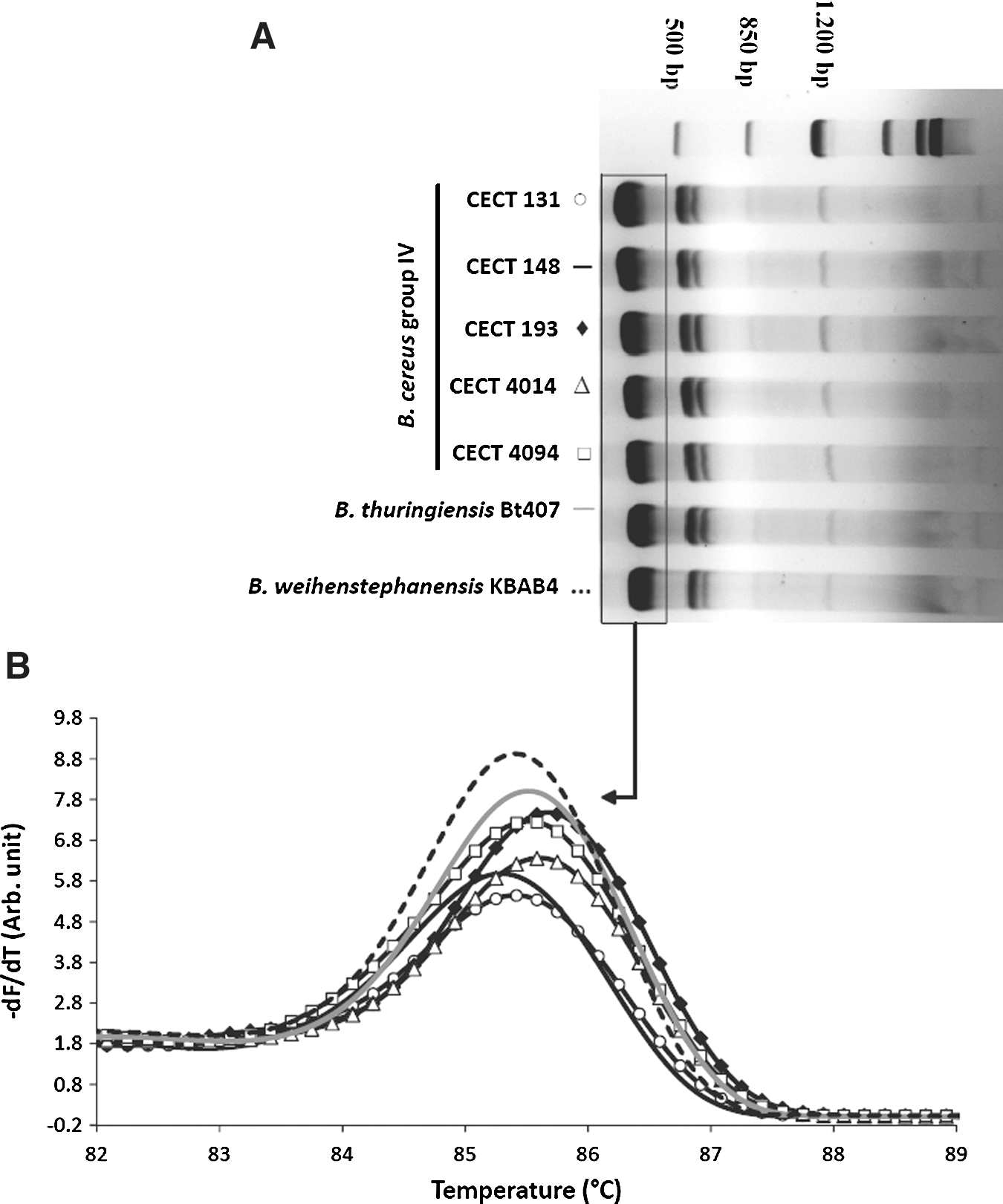
Amplification of 16S-23S intergenic spacer region (ISR) of Bacillus cereus Group strains with primers 16S-23SISRfor and 16S-23SISRrev.
We therefore applied melting peak analysis to 16S-23S ISR, as well as to 5S-GT and16S-ISR and observed two main peaks for 16S-23S ISR PCR product and one peak for 5S-GT and 16S-ISR PCR products, respectively. Tm values were very similar and did not allow consistent differentiation among species and strains. Figure 3B shows the major melting peak of 16S-23S ISR PCR product for all B. cereus sensu stricto strains, B. thuringiensis, and B. weihenstephaniensis.
Evaluation of HRM analysis for species genotyping
B. cereus CECT 148, B. thuringiensis Bt407, and B. weihenstephanensis KBAB4 strains were used to test species discrimination by HRM analysis, following the amplification of polymorphic regions 16S-23S ISR, 5S-GT, and 16S-ISR (Table 2). 5S-GT and 16S-ISR amplicons gave derivative melting curves with a single peak (Fig. 4A,B). 16S-23S ISR PCR products showed two melting peaks. Normalized plots for genotyping were generated by selecting appropriate areas from the melting curves (Fig. 4C,D). Figure 4E,F shows difference plots that were generated by defining representative genotypes followed by subtraction of average plots of the different genotypes from one selected genotype. HRM results indicate that amplicons for regions 5S-GT and 16S-ISR but not for 16S-23S ISR (data not shown) can be applied for discrimination between B. cereus CECT 148, B. thuringiensis Bt407, and B. weihenstephanensis KBAB4 (Fig. 4A–F).

Fluorescence derivative high-resolution melting (HRM) curves of Bacillus cereus Spanish Type Culture Collection (CECT) 148 (black lines), B. thuringiensis Bt407 (gray lines), and B. weihenstephanensis KBAB4 (dashed lines) are shown in
Evaluation of HRM analysis for strain genotyping
We evaluated the potential to discriminate among B. cereus strains based on polymorphism detection through HRM analysis of amplicons 16S-23S ISR, 5S-GT, and 16S-ISR. Melting curves of the 16S-23S ISR region for all the strains tested were characterized by two variant domains, corresponding to the two melting peaks observable in the derivative plot (Fig. 5A). For genotyping with 16S-23S ISR amplicons, a combinatory approach was applied, analyzing the two melting domains separately as well as in combination, because successful genotyping for a given domain varied among the biological replica. Figure 5B,C shows the normalized HRM curve and the difference plot generated for the upper domain 1. PCR products 5S-GT and 16S-ISR did not discriminate between B. cereus strains, and these polymorphic regions were rejected for strain genotyping by HRM analysis.
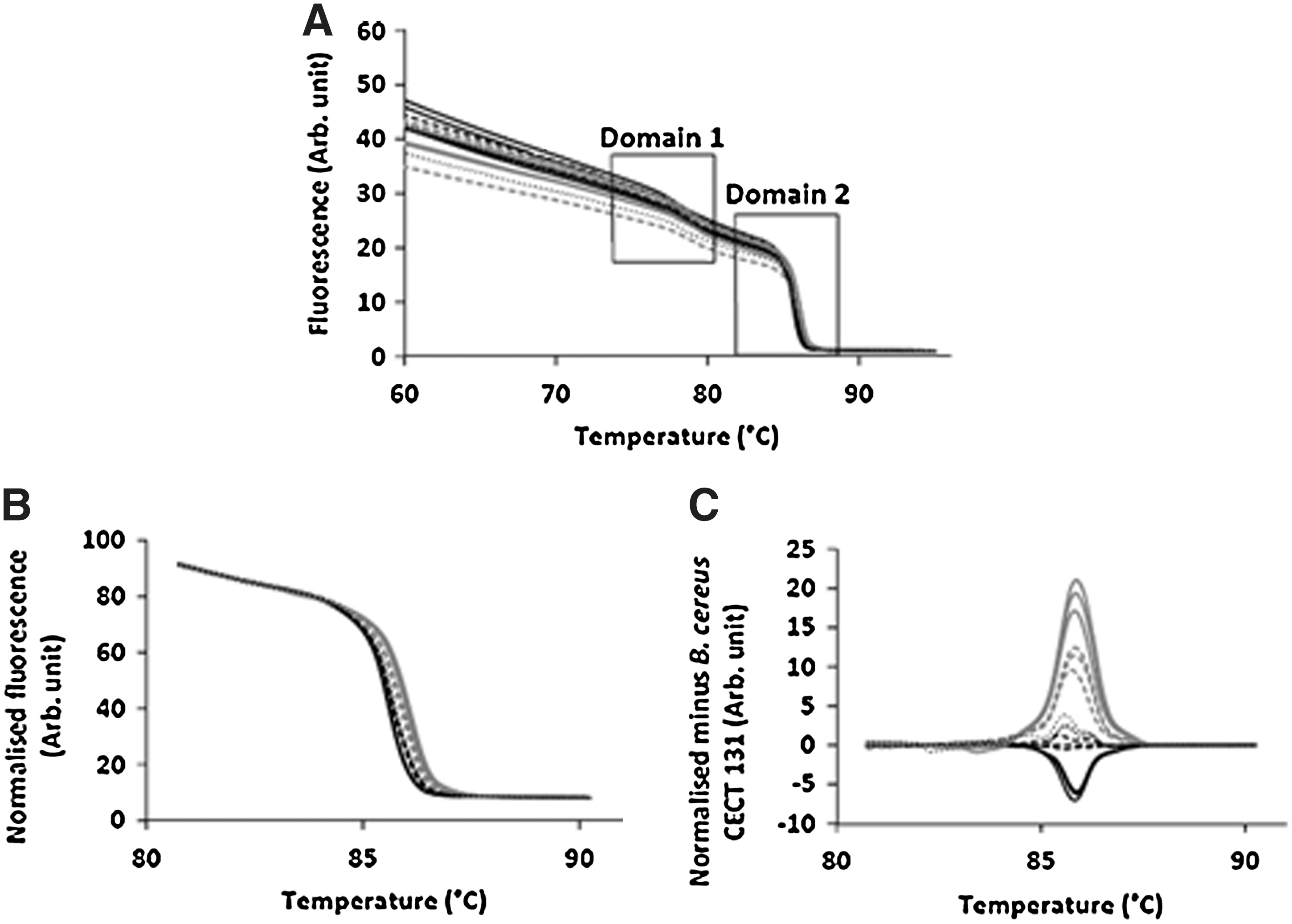
Discussion
Heat resistance values of B. cereus strains connected to diarrheal foodborne outbreaks, food, and environment, and affiliated to group IV, have been reported (Carlin et al., 2006) and were higher than those obtained in this study, indicating variability among group IV strains. Furthermore, K max and shoulder length values obtained also indicate variations within B. cereus CECT strains. Therefore, accurate identification of B. cereus strains could be a valuable tool for food processing.
Phylogenetic analysis of the panC gene sequences showed that CECT B. cereus strains tested are closely related to other strains of the same species as well as B. thuringiensis and B. weihenstephanensis strains. This phylogenetic assessment is consistent with others previously performed (Schmidt et al., 2011).
Genotyping of the 16S-23S ISR has been widely used for differentiation of closely related bacterial species, such as Bacillus genus isolates, based on its high degree of sequence polymorphism (Jensen et al., 1993; Wunschel et al., 1994; Nagpal et al., 1998; Shaver et al., 2002; Cherif et al., 2003). In accordance with Daffonchio et al. (1998, 2000), we observed identical and reproducible amplification patterns in all strains tested, suggesting that the length of 16S-23S ISR is conserved among B. cereus isolates and that different techniques should be used to discriminate between closely related isolates (Martínez-Blanch et al., 2010).
We were not able to distinguish on the species or strain level based on melting peak analysis of 16S-23S ISR using SYBR Green I, probably due to the fact that T m differences of 1°C are not valuable for genotyping and identification (Odell et al., 2005).
On the basis of these results, two primers closely related with rRNA genes and amplifying polymorphic regions, 5S-GT and16S-ISR, were designed for genotyping. Melting peak analysis did not allow to discriminate between B. cereus CECT strains and those from B. weihenstephanensis and B. thuringiensis.
In contrast, genotyping by one-step post-PCR HRM analysis for amplicons 5S-GT and 16S-ISR had a high discriminatory power between B. cereus CECT 148, B. thuringiensis Bt407, and B. weihenstephanensis KBAB4 and an excellent typeability, whereas the selected 16S-23S ISR region was not valuable for species differentiation. Conversely, strain typing using 16S-23S ISR region was shown to be an appropriate method for B. cereus CECT strains discrimination. HRM analysis detected polymorphisms in both melting domains, and we propose a combinatory approach considering both melting domains for genotyping. Our findings led us to consider HRM analysis for genotyping as a valuable tool for B. cereus Group typing; however, additional experiments with a major number of species and strains are required to address this issue.
Conclusion
Heat resistance assays revealed significant differences in thermal inactivation of spores from B. cereus sensu stricto strains assigned to group IV. Thermal processing depends on rapid and reliable identification of strains or species. HRM analysis for genotyping detection proved to be discriminatory among B. cereus species and strains. Our results demonstrate that HRM is a fast and sensitive genotyping technique, capable of detecting sequence changes in only a single base. This finding opens up the possibility of implementing HRM analysis as a rapid discrimination method for B. cereus Group isolates.
Footnotes
Acknowledgments
We thank Luis Pedro García González of SEDIC UPCT service for the excellent technical assistance, José Saura for his help in performing heat resistance assays, and Izaskun Mallona for her invaluable assistance to the present work. This project was funded by Ministerio de Educación y Ciencia Español (AGL 2006-13320C03-02/ALI). V. Antolinos is the recipient of a Ph.D. fellowship from the Ministerio de Ciencia e Innovación Español.
Disclosure Statement
No competing financial interests exist.