
Abstract
Human noroviruses (HuNoVs) are the most frequent cause of foodborne viral gastroenteritis, causing approximately 90% of non-bacterial epidemic outbreaks around the world. Rubus coreanus is a species of black raspberry, rich in polyphenols, and known to exert anti-inflammatory, antibacterial, and antiviral activities. In the present study, the antiviral effects of R. coreanus juice (black raspberry [BRB] juice) on foodborne viral surrogates, murine norovirus-1 (MNV-1) and feline calicivirus-F9 (FCV-F9), were compared with those of cranberry juice, grape juice, and orange juice by plaque assays. Among the four juices tested, BRB juice was the most effective in reducing plaques formation of these viruses. Time-of-addition experiments were designed to determine the mechanism of action of BRB juice on MNV-1 and FCV-F9. The maximal antiviral effect of BRB juice against MNV-1 was observed when it was added to RAW 264.7 cells (mouse leukemic monocyte macrophage cell line) simultaneously with the virus. Pre-treatment of either Crandell Reese Feline Kidney cells or FCV-F9 with BRB juice exhibited significant antiviral activity. The inhibition of viral infection by BRB juice on MNV-1 and FCV-F9 probably occurs at the internalization of virions into the cell or the attachment of the viral surface protein to the cellular receptor. The polyphenol components in BRB (i.e., gallic acid and quercetin), however, did not show any activity against these viruses. Our data provide great promise for the utilization of BRB in the prevention of foodborne viral outbreaks.
Introduction
H
Antiviral activities of natural food materials against diverse viruses have been recently reported. Cranberry juice was shown to inhibit the adsorption of bacteriophage T4 to its bacterial host cells and to prevent the replication of simian rotavirus in its monkey kidney host cells (Lipson et al., 2007a). Cranberry high-molecular-weight proanthocyanidin and grape proanthocyanidin extracts markedly reduced reovirus infectivity titers in cell culture (Lipson et al., 2007b). Other recent studies demonstrated antiviral activity of natural food materials against HuNoV surrogates, MNV-1 and FCV-F9. For example, grape seed extract treatment was found to greatly reduce FCV-F9 and MNV-1 titers (Su and D'Souza, 2011). More than 50% of the total titers of MNV-1 and FCV-F9 were reduced after treatment with cranberry juice (Su et al, 2010).
Rubus coreanus, which belongs to the Rosaceae family, is a species of black raspberry widely known as bokbunja, which is native to Korea, China, and Japan. The fruit of R. coreanus is rich in polyphenols, which are known to have biological activities that may positively impact human health. It has the highest antioxidant capacity among fruits and vegetables (Garcia-Alonso et al., 2004) and has been used in the production of traditional wine and beverages. It was previously shown to possess bone protecting effects (Do et al., 2008), antioxidant activity (Ku and Mun, 2008), and antibacterial activity against foodborne bacterial pathogens such as Escherichia coli O157:H7, Salmonella typhimurium, Bacillus cereus, and Staphylococcus aureus (Park and Jang, 2003). It was also reported to exhibit strong antiviral activity against hepatitis B virus (Kim et al., 2001). However, studies on the antiviral activity of R. coreanus juice (black raspberry [BRB] juice) against foodborne enteric viruses have not been reported. In the present study, the effects of BRB juice on MNV-1 and FCV-F9 were evaluated using an infectious plaque assay, to present the first report of antiviral activity of R. coreanus against these viruses.
Methods
Viruses and cell lines
Feline calicivirus (FCV-F9), RAW 264.7 cells, and Crandell Reese Feline Kidney (CRFK) cells were obtained from American Type Culture Collection (ATCC; Manassas, VA). MNV-1 was kindly provided by Dr. Jai Myung Yang (Sogang University, Seoul, Korea).
Preparation of fruit extracts and determination of total polyphenols
Extracts of R. coreanus (Sununsan Nonghyup, Korea), cranberry, orange, and grape (obtained from a local market) were prepared by hand squeezing the ripe fruit with cheesecloth for 20 min. This was incubated in a shaking water bath at 63°C for 30 min and then centrifuged at 14,000×g for 30 min at 4°C. After centrifugation, the supernatants were collected and sterilized by filtration through 0.22-μm filters. Total polyphenols of juice were determined using the Folin-Ciocalteu method (Singleton et al., 1999). Briefly, the juices were oxidized with Folin-Ciocalteu reagent, and the reaction was neutralized with sodium carbonate. The absorbance was measured at 765 nm using a microplate reader (SpectraMax M2; Molecular Devices Corp. USA, Sunnyvale, CA). Gallic acid was used as a standard total phenolic content. The results were expressed as milligrams of gallic acid equivalent (GAE) per mL.
Cytotoxic assay of BRB juice and polyphenols (gallic acid and quercetin)
RAW 264.7 cells were seeded in 96-well tissue culture plates at a density of 1.5×106 viable cells per mL in Dulbecco's modified Eagle's medium (DMEM) containing 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin (PS). After the cells were incubated at 37°C with 5% CO2 for 24 h, the media were removed. DMEM containing 10% FBS and one of the four different inhibitors—BRB juice (final concentration 3% or 6%), gallic acid, or quercetin—were added to the cells. The cells were further incubated for 1 h at 37°C and 5% CO2. In the case of CRFK cells, a density of 2×105 viable cells per mL was used. The cytotoxicity was measured by an MTT assay; 10 μL MTT, 3-(4,5-dimethylthiozol-2-yl)-3,5-dipheryl tetrazolium bromide, solution was added to each well and incubated at 37°C for 2 h. After removal of supernatant, 100 μL of DMSO was added for solubilization of formazan crystals and incubated for 30 min. The relative absorbance at 570 nm was determined in a microplate reader (SpectraMax M2, Molecular Devices Corp. USA). The percentage of cell viability after treatment with BRB juice was calculated as follows: % cell viability=(treatment / control) ×100.
Cytopathic effect (CPE) reduction assay of polyphenols
Gallic acid and quercetin are main phenolic compounds of fruit of R. coreanus (Ju et al., 2009; Yoon et al., 2003). The confluent cell monolayers were infected with MNV-1 or FCV-F9 in the absence (control) or presence of gallic acid or quercetin. After incubation for 72 h at 37°C and 5% CO2, the virus-induced CPE was analyzed by an MTT assay. The percentage of cell viability was calculated as above, relative to the treatment with virus only. All measurements were performed in triplicate.
Plaque reduction assay of BRB juice
Antiviral activity was evaluated by plaque reduction assays at the indicated multiplicities of infection (MOI). Pre-treatment or co-treatment assessed the ability of BRB juice in preventing the attachment of virus to cells or inhibiting the internalization of virus; post-treatment assays were used to test the ability of preventing the replication of virus (Fig. 1).
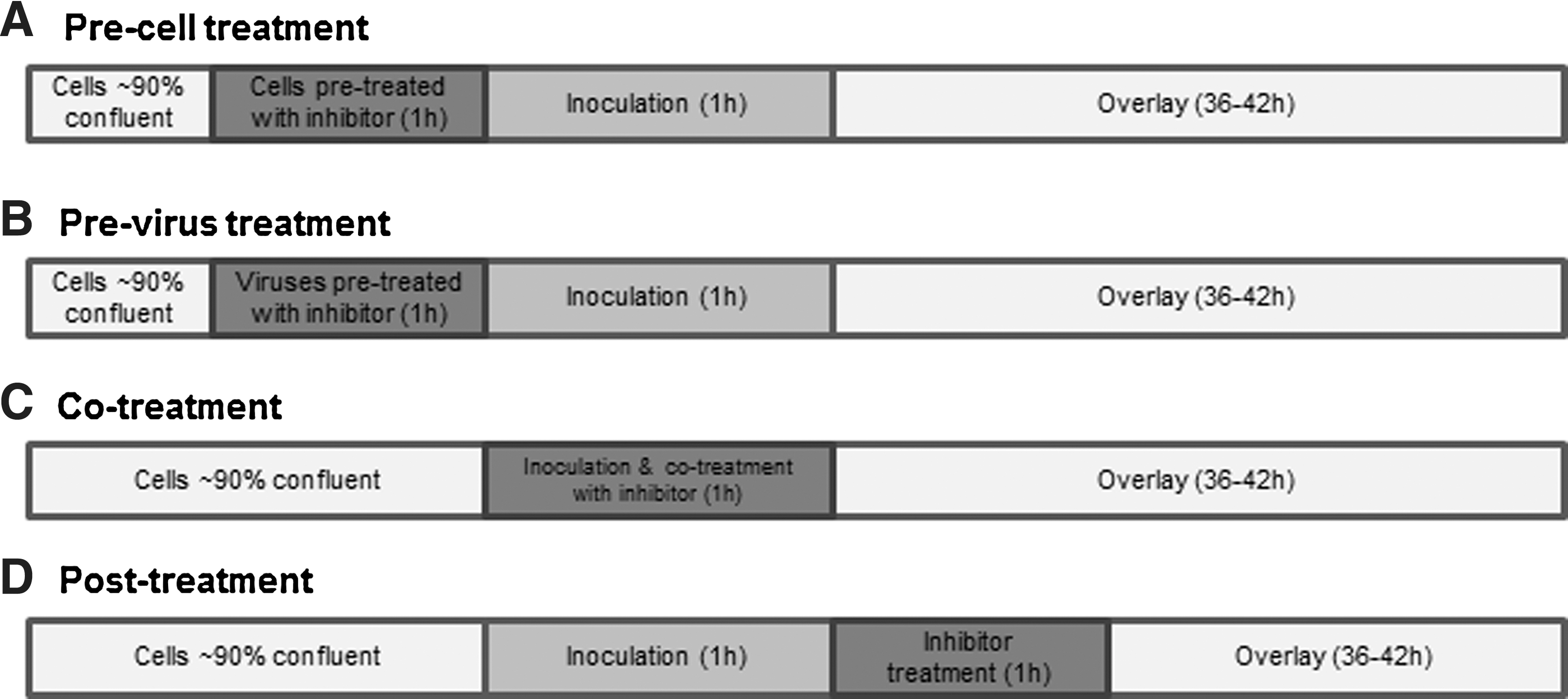
Flow charts of the time-of-addition assay describing different treatment conditions.
Pre-treatment of BRB juice was applied to cells and viruses separately. Pre-treatment of RAW 264.7 cells with BRB juice was conducted as follows: DMEM containing 10% FBS and 1% PS, and BRB juice (final concentrations 3% and 6%) was added to confluent monolayers of RAW 264.7 cells and incubated at 37°C in a CO2 incubator for 1 h with occasional rocking. After complete aspiration of the cell media containing the juice, 10-fold serial dilutions of virus stocks (100 μL) prepared in DMEM containing 10% FBS and 1% PS were inoculated into each well. After viruses were adsorbed for 1 h at 37°C in a CO2 incubator, the inocula were removed and 1 mL of DMEM containing 1.5% agarose, 5% FBS, and 0.5% PS was added to each well. The plates were incubated for 42 h at 37°C in a CO2 incubator; then the cells were stained with 0.5% crystal violet, and plaques were counted afterwards. Sterilized distilled water was used as the untreated control.
Pre-treatment of MNV-1 with BRB juice was performed by mixing equal volumes of the same juice (final concentrations 3% and 6%) and MNV-1 to reach titers of 5×105 PFU/mL and incubated at room temperature for 1 h with occasional rocking. After incubation, treatment was neutralized by 10-fold serial dilutions of virus stock in DMEM containing 10% FBS. The dilutions of virus were inoculated onto confluent RAW 264.7 cell monolayers for 1 h at 37°C in a CO2 incubator. After virus adsorption, the rest of the procedure was as described for the pre-treatment of cells.
In co-treatment of BRB juice, the same procedure as with the pre-treated cells was conducted, except that confluent RAW 264.7 cell monolayers were infected with 100 μL of 10-fold serial dilutions of virus stocks, which was simultaneously mixed with BRB juice (final concentrations 3% and 6%) for 1 h at 37°C in a CO2 incubator. In post-treatment, after virus adsorption to the cells, the inocula were completely removed and the cells were incubated with BRB juice for 1 h; the rest of the procedure was as described for the pre-treatment of cells. FCV-F9 plaque reduction assays using CRFK cells were also performed based on the method as described for MNV-1 and RAW 264.7 cells.
Statistical analysis
Experimental results are expressed as mean±standard deviation (SD). All measurements were performed in triplicate. The data were analyzed using analysis of variance (ANOVA) with SAS software (version 9.2; SAS Institute, Cary, NC), and the means were separated with Duncan's multiple range test. Differences with a value of p<0.05 were considered significant.
Results
Antiviral activity of fruit juice on MNV-1 or FCV-F9
The cytotoxicity effects of BRB juice, cranberry juice, grape juice, and orange juice on RAW 264.7 cells or CRFK cells were not observed at concentrations up to 6% (p<0.05; data not shown). Subsequent assays were carried out at these concentrations, without losing significant visual effects on the viability of the cells. A comparative study of antiviral effects of BRB juice, cranberry juice, grape juice, and orange juice was done using co-treatment against MNV-1 and FCV-F9. Notably, BRB juice and cranberry juice decreased the plaque formation at concentrations of 3% and 6% (Fig. 2A). However, grape juice and orange juice did not show inhibitory effects on these viruses. In both MNV-1 and FCV-F9, the reduction of plaque formation was found to be consistently higher in the treatment of BRB juice compared to that of cranberry juice. Plaque reduction of MNV-1 and FCV-F9 followed the decreasing order of BRB juice>cranberry juice>grape juice=orange juice. Approximately 75.33±6.60% and 92.67±1.89% plaque reduction in MNV-1 (MOI 0.001) was obtained in the presence of 3% and 6% BRB juice, respectively. In the case of FCV-F9 (MOI 0.001), a reduction of 12.90±9.68% and 48.15±6.41% in plaque formation was observed upon incubation with 3% and 6% BRB juice, respectively (Fig. 2B). The antiviral effect of BRB juice was further evaluated at different time points during virus infection to assess the mechanism of antiviral activity (Fig. 1).
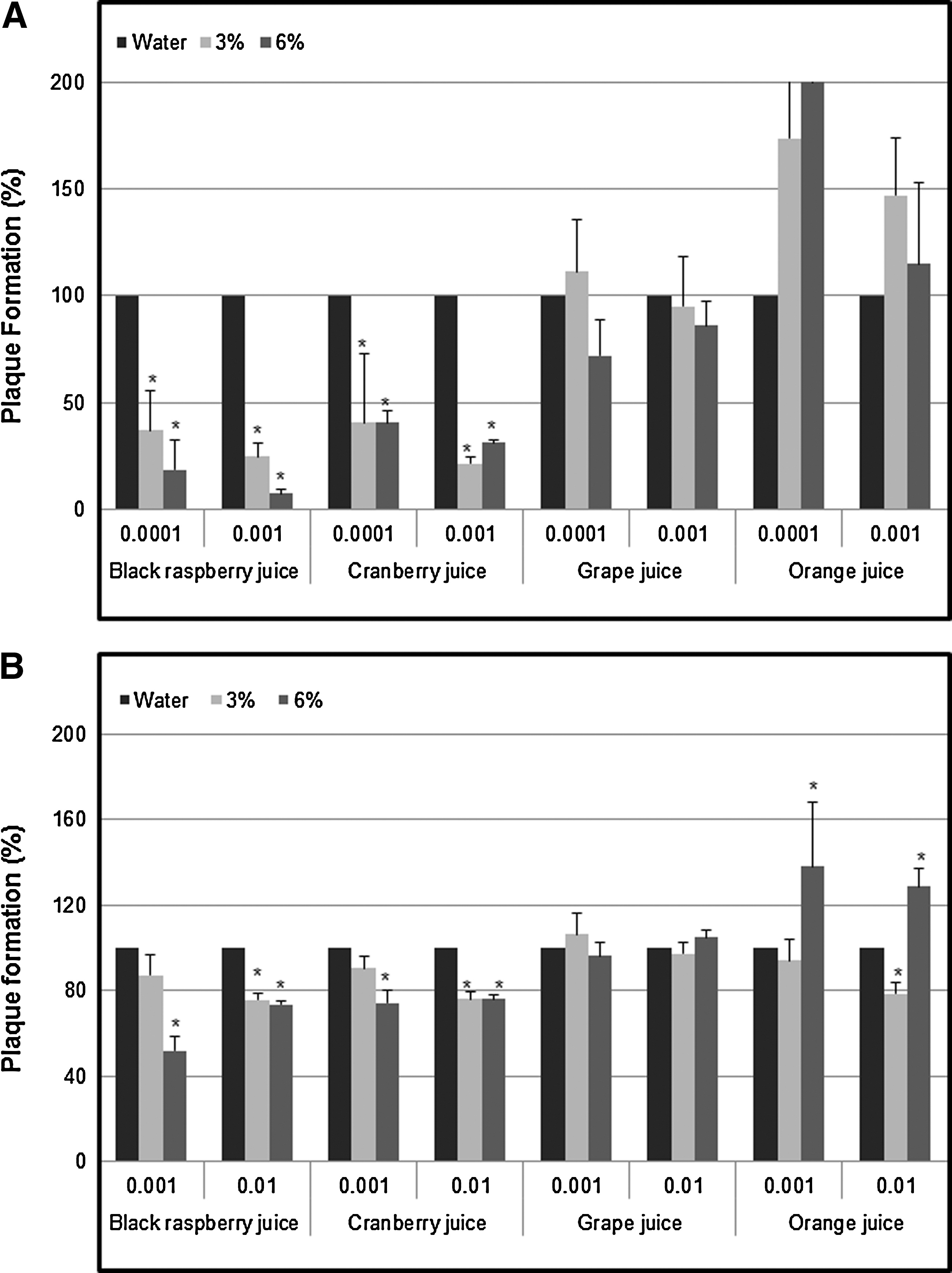
Inhibition of plaque formation by various fruit juices using co-treatment. Cells were infected with murine norovirus-1 (MNV-1)
Antiviral activity of BRB juice on norovirus attachment or internalization
When the cells were treated with BRB juice prior to virus infection, it was found that BRB juice possessed antiviral activity against MNV-1, showing 52.45±3.70% and 59.69±13.62% inhibition at 3% and 6% concentrations at MOI 0.001, respectively (Fig. 3A). Pre-treatment of MNV-1 (MOI 0.001) with BRB juice showed lower antiviral activity (e.g., 31.55±10.55% inhibition at 3% BRB juice). In the case of FCV-F9, pre-treatment of CRFK cells with BRB juice exhibited strong antiviral activity against FCV-F9 (MOI 0.0001), showing 71.43±24.74% and 83.33±28.87% inhibition at 3% and 6% concentrations, respectively. Similarly, pre-treatment of FCV-F9 (MOI 0.0001) with BRB juice showed strong antiviral activity, reaching 100.00±0.00% inhibition at 3% (Fig. 3B).
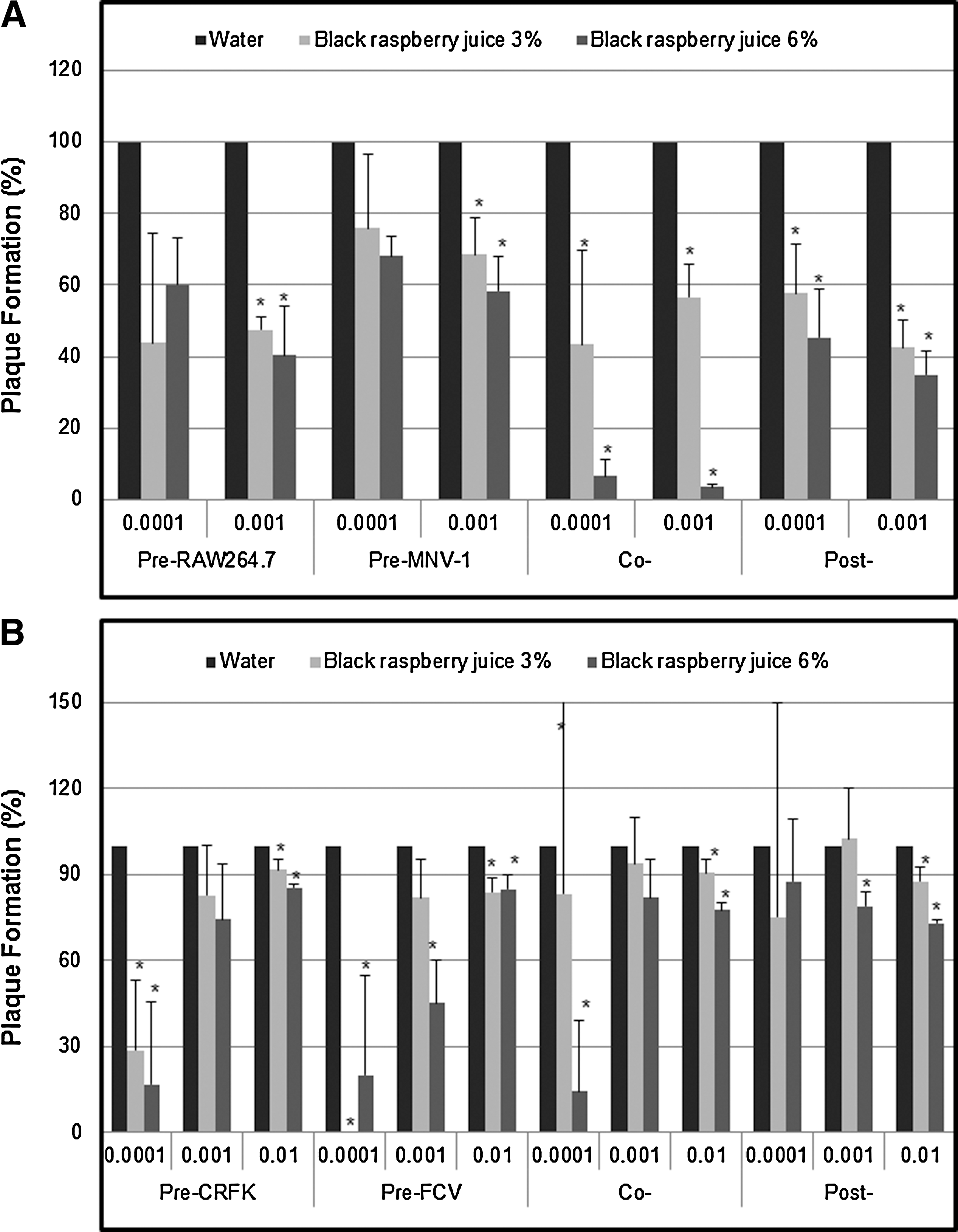
Influence of black raspberry juice on murine norovirus-1 (MNV-1)
In the co-treatment, BRB juice showed a very high antiviral activity against MNV-1 (MOI 0.001), reaching 43.37±9.01% and 96.43±0.72% inhibition at 3% and 6% concentrations, respectively, whereas it showed mild inhibitory activity against FCV-F9 in CRFK cells (Fig. 3A,B). Our results demonstrated that BRB juice had the maximal antiviral activity against MNV-1, when it was used simultaneously with the virus in cells. Also, it exhibited significantly high antiviral activities against FCV-F9 in both cases of pre-treated CRFK cells and pre-treated FCV-F9. These findings suggest that BRB juice can affect MNV-1 and FCV-F9 entry into the cells by blocking binding of the virus to cells, inhibiting the internalization of virus into cells, or by direct effect on either virus particles or binding to cell receptors.
Antiviral activity of BRB juice on norovirus replication
In the post-treatment assay, BRB juice produced 57.50±7.71% and 65.00±6.60% antiviral activity against MNV-1 (MOI 0.001) with 3% and 6% concentrations, respectively (Fig. 3A). In contrast, the same juice showed mild antiviral activity against FCV-F9. At MOI 0.01, 12.3±5.07% and 27.1±1.55% reduction was achieved with 3% and 6% concentrations, respectively (Fig. 3B). FCV-F9 was found to be less sensitive to post-treatment in BRB juice than MNV-1. Taken together, our time-of-addition experiments revealed that the maximal antiviral effects against MNV-1 were achieved when BRB juice was added simultaneously to cells with viruses, whereas pre-treatment of either FCV-F9 or CRFK cells with the juice showed the strongest antiviral activity.
Antiviral activity of gallic acid and quercetin
The total polyphenolic content of BRB juice was found to be lower (0.64±0.01 mg GAE/mL) than that of cranberry juice (Table 1). With regard to the antiviral activity of BRB juice against FCV-F9 and MNV-1, polyphenols can be the most promising candidate. However, gallic acid or quercetin, the main phenolic compounds of BRB juice, did not decrease CPE in RAW 264.7 cells infected with MNV-1 or CRFK cells infected with FCV-F9 (data not shown).
All measurements were performed in triplicate, and data are presented as mean±SD. Within a row, different superscripts denote significant differences using the Duncan's multiple range test (p<0.05).
Discussion
The effects of cranberry juice, grape juice, and pomegranate juice on the infectivity of enteric viruses have been reported (Lipson et al., 2007a; Su and D'Souza, 2011; Su et al., 2011). Commercial juices caused significant reduction in infectivity titer and structural changes in virus particle. When the antiviral effects of BRB juice, cranberry juice, grape juice, and orange juice were compared in this study, BRB juice was the most effective in reducing plaque formation of foodborne enteric virus surrogates. It is notable that the antiviral activity of BRB juice was more potent than that of cranberry juice, which was recently suggested to have a significant antiviral activity against foodborne viral surrogates (Su et al., 2010).
The multiplication of caliciviruses can be divided into the following steps: attachment of the viral protein to the cellular receptor, internalization of the virion in the cell, replication of virus, and release of the mature virion from the cell (Rohayem et al., 2010). When the antiviral activities of BRB juice against MNV-1 and FCV-F9 were examined using time-of-addition experiments, there was a similar inhibition mechanism on MNV-1 and FCV-F9. A significant reduction of the plaque formation of MNV-1 was observed in the co-treatment of virus with BRB juice in a dose-dependent manner. When either the CRFK cell or FCV-F9 was pre-treated with the juice, significant antiviral activity was achieved. These data suggest that the antiviral effect of BRB juice on MNV-1 and FCV-F9 probably occurs at the virus-cell interface mediated via interactions with cell surface receptor or internalization of the virus into the host cells. This observation was in accordance with the previously published report for the inhibition of plaque formation of FCV-F9 in CRFK cells, where enteric viruses pre-incubated with cranberry juice resulted in significant reduction of infectivity titers (Su et al., 2010). It was recently reported that MNV-1 was susceptible to treatment with cranberry juice and pomegranate juice (Su et al., 2010, 2011). In the case of a simian enteric virus, rotavirus SA-11 was changed into anomalous virus-like particles with the treatment of cranberry juice cocktail. The reductions in viral infectivity titers might be explained by an effect on the host cell surface, possibly through an alteration of viral receptor sites (Lipson et al., 2007a).
The polyphenolic content of BRB juice is 0.64±0.01 mg GAEs per mL, which is lower than that of cranberry juice (1.04±0.05 mg; Table 1) as well as commercial cranberry juice, containing 1.7±0.2 mg (Seeram et al., 2008). The bioactive polyphenols such as proanthocyanidins may play a role in reducing the viral infectivity, possibly through an alteration of viral receptor sites. Cranberry high-molecular-weight proanthocyanidins and grape proanthocyanidins extracts markedly reduced reovirus infectivity titers (Lipson et al, 2007b). Also, BRB juice, which is known to be rich in polyphenols, showed significant antiviral activity on MNV-1 and FCV-F9 in the present study. However, gallic acid or quercetin did not exhibit any antiviral activity on either of the viruses based on the CPE reduction assay. These data suggest that the polyphenol compounds can act synergistically on antiviral activity of foodborne viral surrogate or that they may not be solely responsible for the antiviral activities of BRB juice.
MNV-1 is known to be more resistant to pH and heat than FCV-F9 (Cannon et al., 2006). The sensitivity of FCV to low pH was explained by its uptake mechanism via clathrin-mediated endocytosis for infection (Stuart and Brown, 2006), while MNV-1 is internalized by a clathrin-independent pathway (Perry and Wobus, 2010). However, it has been reported that the low pH imparted by cranberry juice is not a significant factor in antivirus activity to rotavirus, bacteriophage, MNV-1, and FCV-F9 (Lipson et al., 2007a; Su et al., 2010). Moreover, it was recently shown that MNV-1 resistance to environmental conditions correlates with the reported enteric outbreaks from fruits, fruit juices, and fresh-cut vegetables (Sanchez et al., 2012). In this context, the titer reduction of the foodborne viral surrogates when simultaneously treated or pre-treated with BRB juice shows great promise for its utilization in the prevention of foodborne viral outbreaks. Potential applications should include use of BRB juice for viral reduction in ready-to-eat foods such as fresh-cut vegetables, juices, or juice-cocktail combinations in institutional food service. It may also be useful for viral reduction in produce washes, in edible coating/films, and in functional food materials for public health by reducing infections in high-risk populations.
Conclusion
The present study demonstrated that BRB juice was significantly more effective in reducing plaque formation of foodborne enteric virus surrogates than cranberry juice, grape juice, and orange juice. BRB juice showed the maximal antiviral effects against MNV-1 in the co-treatment with virus, while pre-treatment with either CRFK cells or FCV-F9 exhibited significantly stronger antiviral activity. Our results suggest that BRB juice can inhibit MNV-1 and FCV-F9 at the viral internalization onto the cell or the attachment to the cellular receptor. Further studies are needed to address the characterization of active compounds in BRB juice and their specific antiviral mechanisms against MNV-1 and FCV-F9.
Footnotes
Acknowledgments
We thank Professor Y.T. Ko at Duksung Women's University for his comments. This research was supported by the Korea Healthcare Technology R&D Project, Ministry of Health, Welfare and Family Affairs, Korea (grant A085119 to K.H.K.) and the National Research Foundation of Korea, funded by the Korean Government (Ministry of Education, Science and Technology; grant NRF-2011-355-C00119 to J.-H.L.).
Disclosure Statement
No competing financial interests exist.