
Abstract
The purpose of this research was to study the effect of different factors on the resistance of Cronobacter sakazakii NCTC 9238 to UV-C light (which includes germicidal ultraviolet light at 254 nm) and to determine whether a combined treatment with heat would produce a synergistic effect for its inactivation. Growth temperature between 10°C and 37°C did not change the UV-C resistance of C. sakazakii. On the contrary, cells in the logarithmic phase of growth were more sensitive to UV-C light than in the stationary phase. The lethality of UV-C was independent of pH (between 3.0 and 7.0) and aw (between 0.94 and >0.99) of the treatment media, but it exponentially decreased with the absorption coefficient (α). When applying a UV-C treatment of 27.1 J/mL at 25°C to C. sakazakii suspended in vegetable soup and apple juice, 1 and 2 log10 cycles of inactivation were barely achieved, respectively. However, the bactericidal effect of UV-C light increased with temperature. The lethality of the combined process was the result of a synergistic effect that was maximum at 52.5°C for apple juice and 55°C for vegetable soup. In conclusion, these results indicate that UV-C efficacy may be influenced by microbial growth conditions and food characteristics, and that its combination with heat may act synergistically against C. sakazakii.
Introduction
C
Ultraviolet (UV) light, emitted by mercury arc-lamps (low or medium pressure), is effective against a variety of viruses, bacteria, and protozoan cysts (Hijnen et al., 2006). The most effective wavelengths are located between 200 and 280 nm (the so-called short-wave UV light, or UV-C light), especially at 254 nm (Bachmann, 1975; Bintsis et al., 2000). Photons of UV-C light are absorbed by DNA nucleoside bases causing the formation of cross-linking photoproducts and preventing cells from reproducing (Billmeyer, 1997; Giese, 1997; Hijnen et al., 2006). The germicidal effect of UV-C light is well established for air and surface decontamination as well as for water disinfection replacing conventional chlorination processes (López-Malo and Palou, 2005). Furthermore, UV-C light holds considerable promise in food processing as an emerging nonthermal method for microbial inactivation in liquid food products without causing major changes to their sensorial properties (Sizer and Balasubramaniam, 1999) and without generating chemical residues (Guerrero-Beltrán and Barbosa-Cánovas, 2004). In 2004, the NACMCF revised the definition of “pasteurization” and included UV irradiation as an alternative to heat for pasteurization purposes (NACMCF, 2004). Particularly, fruit juices have received increasing attention for being susceptible to being treated by UV-C (Franz et al., 2009; Keyser et al., 2008; Oteiza et al., 2010). In fact, in 2000, the FDA approved UV light treatment as an alternative to heat for the pasteurization of fresh juice products (FDA, 2000). Although there are still few studies conducted on irradiated fruit juices (Falguera et al., 2011), in the validation of equipment to ensure the 5-log reduction, Escherichia coli O157:H7 or a surrogate strain (E. coli American Type Culture Collection [ATCC] 25922) is usually used as target pathogen (Franz et al., 2009; Keyser et al., 2008; Wright et al., 2000) due to its implication in severe outbreaks of unpasteurized juice (CDC, 1997). However, target microorganisms for pasteurization processes may change depending on the selected processing technology. According to some authors (Koutchma, 2009), the most UV-C–resistant microorganisms of public health significance have not been fully determined, and this is especially true if we include emerging pathogens such as C. sakazakii. Today, there are limited data in the literature concerning the UV-C resistance of C. sakazakii (Ha and Ha, 2010; Jo et al., 2010; Liu et al., 2012). Liu et al. (2012) stated that although UV radiation of powdered food is still under investigation, it may be feasible to use UV radiation to inactivate C. sakazakii in dry infant formula if the penetration could be improved.
The objective of this work was to study the sensitivity of C. sakazakii to UV-C light. For this purpose, the effect of microbiological factors (strain, growth temperature, and growth phase) and treatment medium characteristics (pH, water activity, absorption coefficient, and composition) were explored. In order to increase UV-C light lethality, the effect of combining it with moderate heating was also studied.
Materials and Methods
Bacterial culture and media
C. sakazakii CECT 858 (ATCC strain 29544) was provided by the Spanish Type Culture Collection (CECT, Valencia, Spain), and NCTC 8155, 9238, and 9529 strains by the National Collection of Type Cultures (NCTC, Salisbury, UK). Cultures were kept frozen at −80°C in cryovials. A broth subculture was prepared by inoculating, with a single colony from an Oh and Kang (OK) agar plate (Vitaltech Ibérica S.L., Barbera Del Valles, Spain), a flask containing 10 mL of sterile Tryptone Soya Broth supplemented with 0.6% of Yeast Extract (TSBYE; Biolife, Milan, Italy). After inoculation, the flask was incubated overnight at 30°C in a rotary shaker (150 rpm). Flasks containing 50 mL of sterile TSBYE were inoculated with the overnight subculture and then incubated under agitation for 24 h at 30°C to reach the stationary phase (approximately 5×109 CFU/mL). Cells in the exponential phase (approximately 5×108 CFU/mL) were obtained after 6 h of incubation at 30°C (OD680=0.3). The influence of cell age was studied by prolonging the incubation time for 48, 72, and 96 h (approximately 6×109, 6.5×109, and 4×109 CFU/mL, respectively).
McIlvaine citrate-phosphate buffers (Dawson et al., 1974) of pH 3.0, 4.0, 5.0, 6.0, and 7.0 (aw>0.99) were used. Sucrose (Azucarera Ebro, Madrid, Spain) and tartrazine (Sigma-Aldrich, St. Louis, MO) were added at different concentrations to pH 7.0 buffer to obtain media of different aw and absorption coefficient (α), respectively (see Table 2 below). Commercially sterilized apple juice (pH 3.4; aw>0.99; α=22.1 cm−1; 5.4 NTU; Alcampo, S.A., Madrid, Spain) and vegetable soup (pH 5.8; aw>0.99; α=23.5 cm−1; 1810 NTU; Interal S.A., Lezo Guipuzcoa, Spain) were also used as treatment media.
Analytical measurements
Media absorbance was measured at 254 nm using a Unicam UV500 spectrophotometer (Unicam Ltd., Cambridge, UK). Sample solutions were diluted and evaluated using quartz cuvettes (Hellma, Müllheim, Germany) with path lengths of 1, 2, and 10 mm. The absorption coefficient (cm−1) of the sample solution was determined from the slope of the absorbance versus the path length correcting the dilution factor. Turbidity (NTU) was measured using a nephelometer (model 83749; Hanna Instrument, Szeged, Hungary), pH using a pHmeter (model Basic20; Crison Instrument, Barcelona, Spain), and aw with a water activity meter (model CX-1; Decagon Devices, Pullman, WA).
UV-C treatments, incubation of treated samples, and survival counting
UV-C treatments were carried out in an apparatus previously described (Gayán et al., 2011), which consists of eight annular thin-film flow-through reactors connected in series. Each reactor is an 8-W low-pressure UV lamp (TUV 8WT5; Philips, Andover, MA) emitting 85% of energy at 254 nm. The circuit and reactors are submerged in a 90-L water bath (Ta±1.5°C) heated by circulating water from a thermostatic bath (model Kattebad K12; Huber, Offenburg, Germany). Two thermocouples (model ZA020-FS; Almeco, Bernburg, Germany) are fitted to the input of the first and to the outlet of the last reactor to control the temperature.
Once 10 mL of the pure culture was added to the 1 L of treatment medium (final cell concentration of approximately 6×107 CFU/mL for stationary-phase cells and 5×106 CFU/mL for exponential-phase cells), it was pumped with a constant flow of 8.5 L/h through the heat exchanger to the reactors in a single pass. When treatment conditions were stabilized, samples were aseptically withdrawn from the flow stream through the sampling valves at the outlet of each reactor, and immediately pour-plated into Triptone Soya Agar supplemented with 0.6% of Yeast Extract (TSAYE; Biolife) and then incubated for 24 h at 35°C. After incubation, CFUs were counted with an Image Analyzer Automatic Colony Counter (Protos; Synoptics, Cambridge, UK) (Condón et al., 1996).
Curve fitting, resistance parameters, and statistical analyses
Survival curves to UV-C treatments were obtained by plotting the log10 fraction of survivors versus the treatment dose (J/mL). Survival curves were fitted to the log-linear regression plus shoulder model (Geeraerd et al., 2000) from the GInaFiT fitting tool (Geeraerd et al., 2005):
where Sl is the shoulder length or dose before the exponential inactivation begins, and kmax is the inactivation rate or the slope of the exponential portion of the survival curve. For comparison purposes, the GInaFiT also provides the 4D value or dose necessary to inactivate 99.99% of the microbial population. In this work, the dose has been calculated using the experimentally calculated average residence time (Gayán et al., 2011).
Statistical analysis
Analysis of variance (ANOVA) tests followed by Tukey's test (p=0.05) were carried out using the GraphPad PRISM 4.1 software (GraphPad Software, Inc., San Diego, CA), and differences were considered significant for p≤0.05. Resistance determinations were performed at least three times in independent experiments. The error bars in the figures correspond to the mean standard deviation (SD).
Results and Discussion
The major factors affecting the lethality of UV-C treatments include the design of the equipment, the type and physiological state of the bacterial cells, and the physico-chemical characteristics of the treatment medium (Franz et al., 2009; López-Malo and Palou, 2005). Regarding the design of the equipment, the main drawback for the use of UV-C light pasteurization of liquid food products is its low penetration capacity (Sizer and Balasubramaniam, 1999). To improve the effectiveness of UV pasteurization equipments, two strategies are being employed: the use of laminar flow thin film UV reactors, and flowing the liquid in a turbulent regime to ensure that all of the liquid is targeted equally by the UV-C light (Koutchma et al., 2004). Regardless of the strategy followed, the problem is the accurate measurement of the UV-C dose because it is the result of multiplying the distribution of light irradiance by the exposure time (Koutchma and Parisi, 2004), which is also a distribution. Therefore, a quantitative comparison of inactivation data obtained with different equipment should be made with caution. The characterization of the equipment used here has been previously published (Gayán et al., 2011) and our data can be directly compared with those of Gayán et al. (2011).
Inactivation kinetics and intraspecific variation in resistance
The UV-C resistance of four C. sakazakii strains was studied in the reference conditions: at 25°C in buffer of pH 7.0, aw>0.99, and α=10.51 cm−1. Figure 1 shows the survival curves of the four strains and depicts the profile of all curves obtained in this work: concave downwards due to the presence of shoulders. This profile can be explained by the following phenomenon. The lethal effect of UV irradiation depends on the number of cross-linking between neighboring pyrimidine nucleoside bases (DNA damage), which is proportional to the amount of UV exposure, and on the damage repair capability of the cells, which depends on the UV repair system present in the target microorganism and the recovery conditions (Liltved and Cripps, 1999; Miller et al., 1999; Yajima et al., 1995). Once the threshold of cross-linking has been exceeded, the number of cross-links is beyond repair, and cell death occurs (Miller et al., 1999). This phenomenon is then reflected in the shape of the inactivation curve: non-microbial inactivation (lag region) at low UV doses followed by a normal log-linear relationship at higher UV doses (Hoyer, 1998; Tran and Farid, 2004).

Inactivation kinetics of Cronobacter sakazakii Spanish Type Culture Collection (CECT) 858 (•), National Collection of Type Cultures (NCTC) 8155 (♦), NCTC 9238 (▾), and NCTC 9529 (▴) under UV-C light treatments (8×8 W, 8.5 L/h) at 25°C in McIlvaine pH 7.0 buffer and absorption coefficient of 10.51 cm−1.
The fitting of the survival curves to the Geeraerd et al. (2000) model (Eq. 1) allowed to calculate Sl values of 6.29, 5.83, 5.88, and 5.78 J/mL, kmax values of 0.59, 0.64, 0.67, and 0.88 mL/J, and 4D values of 21.9, 20.6, 19.7, and 16.3 J/mL for C. sakazakii NCTC 9238, 8155, 9529, and CECT 858, respectively. There were no statistically significant differences among Sl values for the four strains (p>0.05), which seems to indicate that the four strains may have the same repair capability. Conversely, only for CECT 858 strain, significantly higher kmax and lower 4D values were found (p≤0.05). Consequently, C. sakazakii CECT 858 was the most sensitive strain.
There are few data in the literature regarding the UV resistance variability among strains. Sommer et al. (2000), working with seven strains of E. coli, found that the fluence needed to reach 90% inactivation varied by a factor of 10, and Oteiza et al. (2005) found that the decimal reduction times of five strains of E. coli barely changed 85%. Gayán et al. (2011), working under our experimental conditions, found that the 4D value for the inactivation of five E. coli strains changed by 1.5 times, a similar magnitude to that found here with C. sakazakii (4D×1.4). A further comparison of resistance between E. coli and C. sakazakii indicates that the latter is more UV-C resistant: the dose necessary to inactivate 4 log10 cycles of the most resistant E. coli (CECT 4201) and C. sakazakii (NCTC 9238) strains was 16.6 and 21.9 J/mL, respectively. The production of a yellow pigment by C. sakazakii cells (Caubilla-Barron and Forsythe, 2007; Johler et al., 2010) could be associated with its higher UV light tolerance, because yellow pigments usually interact with radiations in the UV range.
Subsequent studies in this work were performed with C. sakazakii NCTC 9238 strain.
Influence of growth conditions: growth temperature and incubation time
Growth temperature is an important factor modifying bacterial cell membrane composition and physiology (Neidhardt et al., 1990). As it has been suggested that photons can interact with cell envelopes components (Montgomery, 1985), growth temperature might be an important factor influencing UV cell resistance. Figure 2 illustrates the log10 cycles of inactivation of C. sakazakii NCTC 9238 cells grown at 10°C, 20°C, 30°C, and 37°C for 140, 48, 24, and 24 h, respectively, after a UV-C treatment of 20 J/mL (3-min treatment at 8.5 L/h). Table 1 also includes the parameters of the growth curves fitted to the Baranyi growth model (Baranyi and Roberts, 1994; Baranyi, 1997) (Eq. 2 in Table 1). Although, as usual, the lag phase decreased and the growth rate increased as the growth temperature was raised (by 10 times when the incubation temperature changed 21.3°C), growth temperature did not affect C. sakazakii sensitivity to UV-C light: neither the Sl nor the kmax values changed with growth temperature (p>0.05; data not shown). To the best of our knowledge, there are no data in the literature addressing the influence of growth temperature on microbial sensitivity to UV-C light to which our results can be compared.
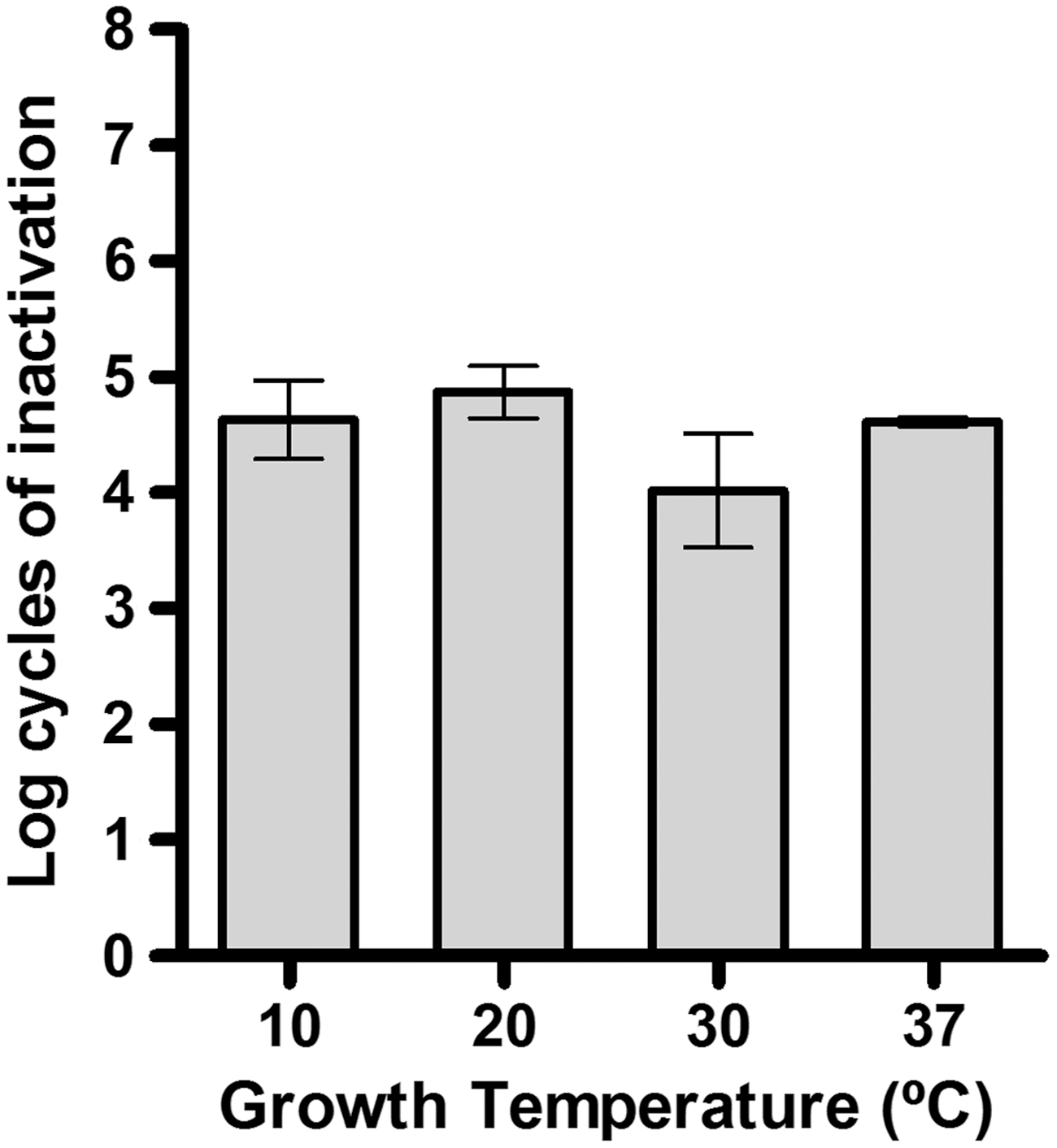
Log10 cycles of inactivation of Cronobacter sakazakii National Collection of Type Cultures (NCTC) 9238 stationary growth-phase cells grown at different temperatures for different times (10, 20, 30, and 37°C for 140, 48, 24, and 24 h, respectively) after 20 J/mL of UV-C light treatment at 25°C in McIlvaine pH 7.0 buffer and absorption coefficient of 10.51 cm−1.
Baranyi growth model (Baranyi and Roberts, 1994; Baranyi, 1997):
y 0, logarithm of the initial cell concentration; y max, logarithm of the asymptotic cell concentration; μ max, maximum specific growth rate; λ, lag time; m and n, curvature parameters; h 0, m×λ; R 2, determination coefficient.
The effect of the incubation time of cells grown at 30°C was also explored (Fig. 3). Results demonstrated that C. sakazakii cells increase their UV resistance when attaining the stationary growth phase, which has been related with the expression of the global stress response gen, rpoS (Bucheli-Witschel et al., 2010). Furthermore, longer incubation times up to 72 h did not significantly (p>0.05) change this maximum value.
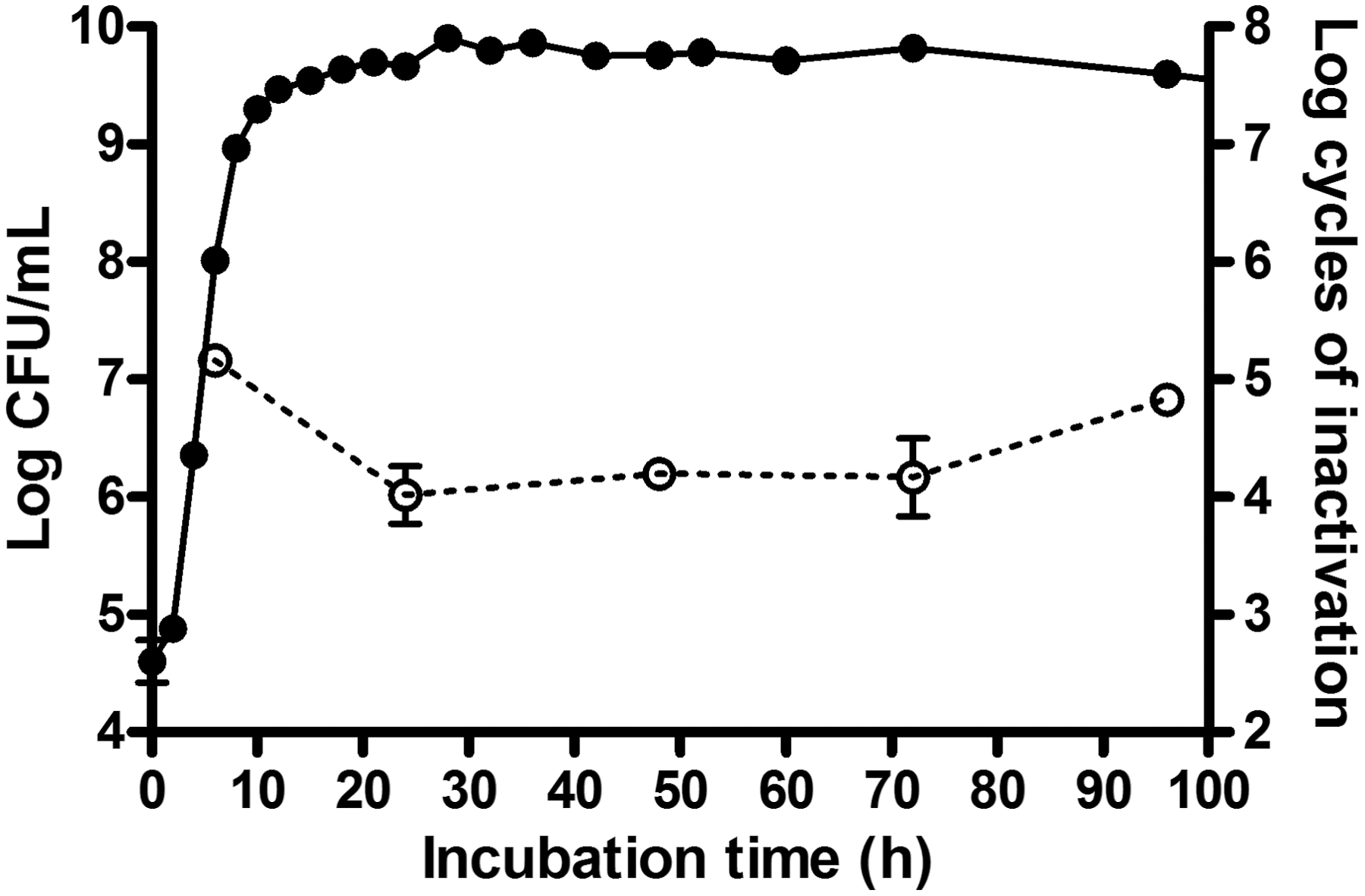
Influence of incubation time on Cronobacter sakazakii National Collection of Type Cultures (NCTC) 9238 UV-C light resistance.
Influence of treatment medium characteristics: pH, aw, and α
It is well known that the physico-chemical characteristics of the treatment medium may change the bactericidal efficacy of most food preservation technologies (Black et al., 2011; Gould, 1992; Stringer et al., 2000). In general, the pH and aw of the treatment medium are the most significant factors, but for UV-C technology, the absorption coefficient is indeed a very important parameter. Table 2 includes the resistance parameters (Sl and kmax ) of C. sakazakii cells in buffer of different pH, aw, and α. Data demonstrated that the lethality of UV-C treatments was independent of pH and aw in the range explored (p>0.05). These results are in agreement with other authors working with other species (Basaran et al., 2004; Gayán et al., 2011; Koutchma et al., 2004; Murakami et al., 2006; Ngadi et al., 2003; Quintero-Ramos et al., 2004) and implies an important distinct characteristic of UV-C technology when compared, for instance, with heat or high hydrostatic pressure (HHP) technologies (Arroyo et al., 2009, 2011).
Analysis of variance (ANOVA) test was conducted within each k max group, as shown in the significance level column. To identify intergroup differences after significant differences in the ANOVA test (p=0.05), multiple comparisons were performed using Tukey's test. Values with the same superscript did not show statistically significant differences (p>0.05).
Sl, shoulder length; k max, rate of inactivation; 4D, time for four log cycles reduction; SD, standard deviation; ns, no significant (p>0.05); *, extrapolated value; RMSE, root mean standard error; R 2, determination coefficient.
On the contrary, efficacy of UV-C light was highly influenced by the absorption coefficient (Table 2). According to the Beer-Lambert-Bouger law, the greater the absorption coefficient is, the lower the penetration of UV-C light will be. If UV-C light is absorbed, it is no longer available for inactivating microorganisms and lower lethal effects will be achieved. As an example, an UV-C treatment of 6.8 J/mL inactivated 6 log10 cycles of the C. sakazakii population in media of α=4.53 cm−1, but barely changed the counts in media of α=20.54 cm−1. Although the effect of the media absorptivity on the lethality of UV light is well known, its effect on the microbial inactivation kinetics has been less well-studied. Some authors (Kouchma et al., 2004; Oteiza et al., 2005) proposed a linear relationship between the decimal reduction dose and the absorptivity of the treatment media. Later on, Gayán et al. (2011) proposed an exponential relationship in a wider range of absorptivities. Our results (Fig. 4) also demonstrate that there was an exponential relationship between kmax
and α for C. sakazakii UV-C inactivation:
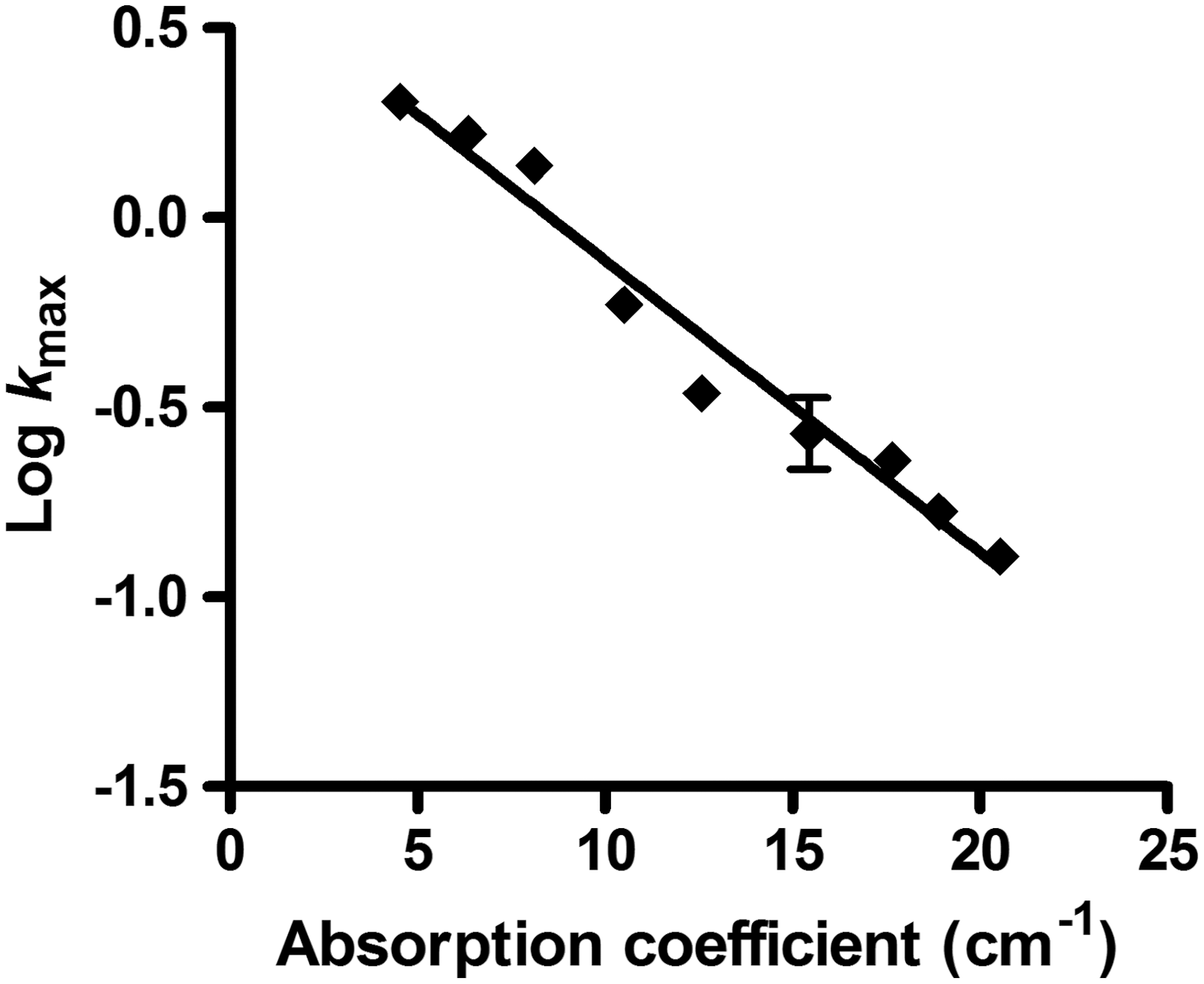
Relationship between the absorption coefficient and log kmax obtained for Cronobacter sakazakii National Collection of Type Cultures (NCTC) 9238 cells (grown at 30°C for 24 h) treated by UV-C light at 25°C in McIlvaine pH 7.0 buffer of different absorption coefficients.
The slope of the regression line that defines this relationship indicates that the inactivation rate decreases by 10-fold when increasing the absorption coefficient by 13 cm−1 units (standard error [SE]=0.57). This value significantly differs (p≤0.05) from that obtained by Gayán et al. (2011) with E. coli (15.92 cm−1), which is an important fact that should be studied in depth with other species.
UV-C resistance in food: Synergistic effect for the combination with moderate heating
Most liquid food products present high absorption coefficients and turbidity, which limit the penetration capacity of the radiation, impairing the use of UV-C light treatments for pasteurization purposes. When applying the most intense UV-C treatment at 25°C at a constant flow rate of 8.5 L/h (27.1 J/mL) to C. sakazakii cells suspended in vegetable soup and apple juice, 1 and 2 log10 cycles of inactivation were barely achieved. Most new methods of bacterial inactivation usually demonstrate shortcomings for food preservation purposes, and some authors have proposed a design involving combined processes of lower intensity but with equivalent or even higher degrees of lethality. Our research group have recently demonstrated that UV-C lethal effect in high absorption coefficient buffers quickly increased with temperature (Gayán et al., 2011). Therefore, the possibility of combining UV-C light with mid-range temperatures to achieve a higher degree of C. sakazakii inactivation in both food products was studied. C. sakazakii inactivation in vegetable soup and apple juice was explored at 50°C, 52.5°C, 55°C, and 57.5°C for a UV-C treatment dose of 12.8 J/mL (2-min treatment at 8.5 L/h). Results showed that the lethality of the combined process was the result of a synergistic effect and its magnitude is shown in Figure 5. The synergistic effect was calculated at each temperature with the following equation:

Percentage of synergistic effect for the inactivation of Cronobacter sakazakii National Collection of Type Cultures (NCTC) 9238 cells (grown at 30 °C for 24 h) suspended in apple juice (•) and vegetable soup (▪) after 2 min of the combined process UV-C light + heat at different temperatures.
The synergistic effect increased with temperature up to a maximum, above which it decreased as a consequence of the higher inactivation rate of heat. The maximum synergistic effect was observed at 52.5°C in apple juice and 55°C in vegetable soup (Fig. 5). Figure 6 shows the C. sakazakii survival curves of the combined process at the optimum treatment condition for each food (at the temperature of maximum synergy). The survival curves to UV-C light at 25°C, and to heat at 52.5°C in apple juice (Fig. 6A) or 55°C in vegetable soup (Fig. 6B) have also been included for comparison. These graphs reveal a process criterion of 22 and 29 J/mL UV-C light dose for the inactivation of 5 log10 cycles of C. sakazakii population in apple juice at 52.5°C and in vegetable soup at 55°C, respectively. Therefore, it is possible to reach 5 log10 reductions in the number of survivors with a low UV-C energetic consumption even in food products with high absorption coefficient and turbidity.
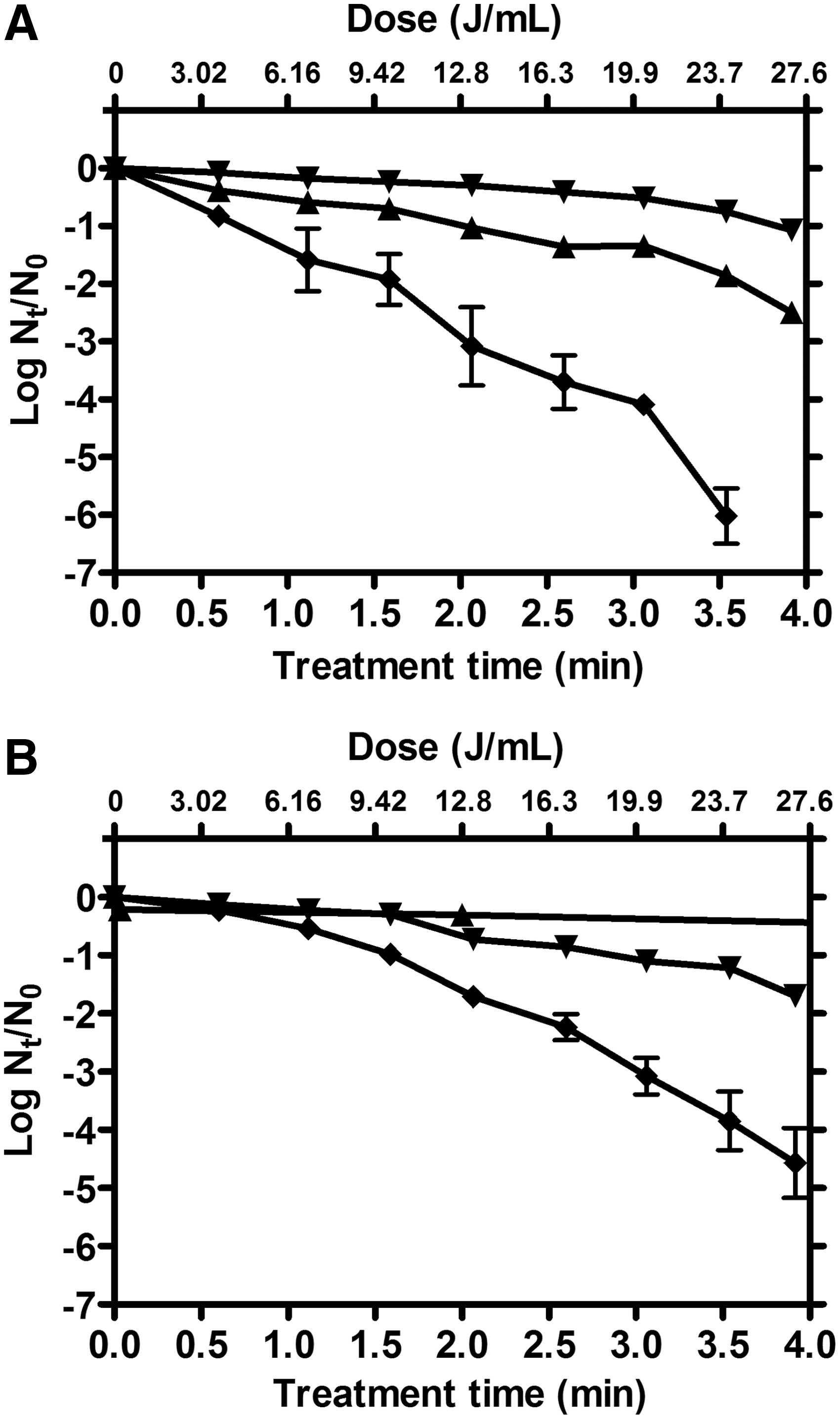
Survival curves of Cronobacter sakazakii National Collection of Type Cultures (NCTC) 9238 cells (grown at 30°C for 24 h) inoculated in apple juice
In conclusion, C. sakazakii resistance to UV-C light is strain-dependent. Its inactivation is influenced by the incubation time and the absorption coefficient of the treatment medium, but not by the growth temperature, and pH or aw of the treatment medium. UV-C light in combination with heat act synergistically to effectively inactivate C. sakazakii population.
Footnotes
Acknowledgments
We thank the Ministerio de Ciencia e Innovación and EU-FEDER (CIT020000-2009-40) for financial support. C.A. and E.G. also gratefully acknowledge the financial support for their Ph.D. work from Gobierno de Aragón and Ministerio de Educación y Ciencia (Spain).
Disclosure Statement
No competing financial interests exist.