
Abstract
There is concern that a relationship exists between antibiotic use in livestock production and the emergence, spread, and persistence of antibiotic resistance. It is important to understand the impact that therapeutic doses of antibiotics for treatment of disease have on resistance because disease treatment typically involves higher doses of antibiotic over short time spans. Absolute quantities of the antibiotic resistance gene bla CMY-2 were measured by real-time quantitative polymerase chain reaction (qPCR) in the bacterial community DNA of fecal samples from dairy cattle that were given a 5-day course of ceftiofur and untreated cattle during a longitudinal, observational study. A hierarchical linear model that accounts for left-censored data and repeated measures was used to estimate group means of bla CMY-2 from the qPCR data. Ceftiofur-treated animals had significantly higher mean quantities of bla CMY-2 than untreated animals during treatment. On the first day post-treatment, mean quantities of bla CMY-2 returned to pre-treatment levels and remained low in both groups for the remainder of the study. The use of qPCR to measure bla CMY-2 quantities provided evidence that the burden of resistance in treated animals may have increased temporarily, a result that was not evident when using only cultivation-based methods of testing for resistance.
Introduction
T
Third- and fourth-generation cephalosporins are one class of antimicrobials that have been identified as critical to both human and animal health (FAO, 2008). Ceftiofur is the only third-generation cephalosporin that is licensed in the United States for use in livestock. In cattle, ceftiofur is currently labeled for the treatment of bovine pneumonia, interdigital necrobacillosis, acute metritis (in cows 0–14 days postpartum), and mastitis, as well as the control of respiratory disease (Pfizer Animal Health, 2011). All administrations of ceftiofur are by prescription of a licensed veterinarian only. Bacterial isolates that are resistant to ceftiofur can be cross-resistant to ceftriaxone, a third-generation cephalosporin that has a variety of applications in human medicine and is the primary choice of treatment of invasive salmonellosis in children (Hohmann, 2001). Because of the increasing resistance to ceftiofur in some bacteria and evidence of cross-resistance with other important antibiotics (CDC, 2012), the U.S. Food and Drug Administration restricted extra-label uses of the cephalosporin class in animal agriculture effective April 5, 2012 (FDA, 2012).
Studies that have investigated the relationship between ceftiofur use and resistance in intestinal microflora of cattle have often focused on generic Escherichia coli because it is easily isolated from bovine fecal samples and frequently possesses multidrug (MDR) plasmids bearing cephalosporin resistance genes (Call et al., 2010; Donaldson et al., 2006; Winokur et al., 2001). These studies have produced mixed results. Cross-sectional studies of dairy herds have reported an association between ceftiofur use at the herd level and ceftiofur-resistant E. coli (Tragesser et al., 2006), while others have reported no relationship (Daniels et al., 2009; Heider et al., 2009). However, the study that reported an association between prior ceftiofur use and ceftiofur-resistant E. coli at the herd level observed no association at the individual animal level (Tragesser et al., 2006). Experimental studies have reported higher levels of ceftiofur-resistant E. coli in ceftiofur-treated cattle compared to untreated cattle that persisted at least 17 days in dairy calves or up to 20 days in adult beef cattle (Jiang et al., 2006; Lowrance et al., 2007). An observational study that followed diseased dairy cattle treated with ceftiofur and healthy, untreated dairy cattle reported no difference between treatment groups in the proportion of E. coli isolates with reduced susceptibility to ceftiofur (Mann et al., 2011).
Among the genes that have been identified as encoding proteins that convey resistance to cephalosporins, bla CMY-2, a β-lactamase encoding gene, is commonly associated with resistant Salmonella and E. coli isolates (Allen and Poppe, 2002; Poole, 2004; Winokur et al., 2000; Zaidi et al., 2007). The bla CMY-2 gene has been identified in a variety of diverse members of the Enterobacteriaceae species and is primarily known as a plasmid-encoded gene (Barlow and Hall, 2002; Call et al., 2010; Jacoby, 2009; Poole, 2004). Given this gene's ability to spread via horizontal gene transfer, the relationship between ceftiofur use and resistance may be better understood by studying bla CMY-2 in the entire bacterial population of a sample rather than particular bacterial species. One study used quantitative real-time polymerase chain reaction (qPCR) to measure bla CMY-2 quantities in the community bacterial DNA of fecal samples from feedlot cattle that received three different treatment regimens of ceftiofur (Alali et al., 2009). The group means of bla CMY-2 quantity were higher in treated cattle than in untreated control cattle for two of the treatment regimens. The durations of elevated bla CMY-2 quantities for those two treatment regimens were approximately 1 week following the cessation of treatment.
Previously, we conducted an observational study on one dairy where diseased dairy cattle treated with ceftiofur and healthy, untreated dairy cattle that commingled were followed over time (Singer et al., 2008). The dairy has a single large barn in which all 150 cows live. All cows can commingle except when they are walked to the milking facility twice a day. Ceftiofur-resistant E. coli could be isolated from the treatment group during treatment and shortly after treatment ended, but no ceftiofur-resistant E. coli were isolated from the untreated group on any day of the study. The resistant bacterial population in the treated cows did not increase in quantity, and the ceftiofur-resistant isolates could no longer be detected by 5 days post-treatment. The bla CMY-2 gene was detected by PCR in samples from both untreated and treated cattle on every day of the study. These results led the authors to conclude that the determinants of ceftiofur resistance were always present in the bacterial populations of these dairy cattle and that ceftiofur treatment temporarily reduced the susceptible bacterial population sufficiently so that ceftiofur-resistant E. coli could be isolated.
The objectives of the current study were to use community DNA extraction and qPCR to quantify bla CMY-2 in the feces of these dairy cattle. We hypothesized that short-term therapeutic treatment would result in a temporary increase in bla CMY-2 quantity during and shortly after treatment but that the average bla CMY-2 quantity would not differ between treated and untreated animals as the time post-treatment increased. Furthermore, we aimed to utilize a novel approach to analyzing qPCR data when gene copy number in individual reactions is low. This study extends the potential utility of qPCR as a more sensitive approach for evaluating the strength of selection pressures on resistant bacterial populations and highlights a more unbiased statistical approach for evaluating this relationship.
Materials and Methods
Study design
This study used the samples obtained from an observational investigation of dairy cattle that were treated with ceftiofur for infertility due to Leptospira borgpetersenii serovar Hardjo-bovis, as previously described (Singer et al., 2008). Following the veterinarian's recommendation, five diseased cattle were treated according to label instructions (2.2 mg/kg, intramuscularly, once daily for 5 days), and all five treated cows started therapy at the same time. Fecal samples were collected from the five treated cows and five untreated animals prior to (days 0 and 1), during (days 3 and 5), and after (days 6 to 12, 15, and 26) ceftiofur therapy. On day 1, sample collection occurred approximately 1 h before the first ceftiofur injection was administered. DNA was extracted from each fecal sample as previously described (Patterson and Singer, 2006; Singer et al., 2008).
Quantitative real-time PCR
Standard preparation
For absolute quantification of bla CMY-2 in fecal samples, plasmid standards were created by cloning a single copy of the bla CMY-2 amplicon into the pDrive cloning vector (PCR Cloning Kkit; Qiagen, Valencia, CA). Plasmid DNA containing the cloned gene was purified using a QIAprep Spin Miniprep Kit. The purified plasmid was checked by PCR using SP6 and T7 primers to assure the gene was present as a single copy. The purified plasmid was quantified by A260 and the copy number per μL was calculated based on the size of the plasmid plus the insert. Aliquots of the standards, diluted in nuclease free water to concentrations of 1×1010 copies per 5 μL, were stored at −20°C. For each qPCR experiment, a new aliquot was thawed and 10-fold serial dilutions were made in a 1:25 mixture of BSA and nuclease free water down to 1×101 copies per 5 μL. Addition of BSA to the dilutions was found to improve the sensitivity of the assay.
Real-time PCR reaction and quantification
We designed, optimized, and validated with nucleotide sequencing the primers 5′-CAG CAT CTC CCA GCC TAA TC-3′, and 5′-GAA GCC CGT ACA CGT TTC TC-3′. Primers were designed with Primer3 software (Rozen and Skaletsky, 2000) using the sequences of the plasmid-borne bla CMY-2 gene in E. coli. QPCR reactions were performed on the unknowns on a Strategene Mx3000p system (Life Technologies Corp., Carlsbad, CA) using Brilliant® II SYBR® Green qPCR mix according to the following recipe: 12.5 μL SYBR Green master mix, 0.5 μL (300nM) of each primer, 1 μL of BSA, 0.375 μL of ROX reference dye (1:500 dilution in nuclease free water), 5.125 μL of nuclease free water, and 5 μL of plasmid standard or unknown community DNA for a total volume of 25 μL per QPCR reaction. The master mix for the plasmid standards followed the same recipe as the unknowns with the exception that water was substituted for the BSA, which had been previously added to the standards during the serial dilution step. The reaction conditions for the amplification were 95°C for 10 min and 40 cycles of 94°C for 30 sec 60°C for 1 min and 72°C for 30 sec each. Fluorescence was measured at the end of the extension phase of each cycle. Following the last cycle, a continuous dissociation curve for each sample was created by measuring fluorescence at short temperature intervals ranging from 60°C to 95°C. Standards and no template controls were included with all of the assays and all standards, unknowns, and controls were run in triplicate.
Quantification of blaCMY-2 in unknown samples
The dissociation curves for all samples were examined to confirm that only one peak at the correct melt temperature was present, providing evidence that no non-specific amplification occurred. Gel electrophoresis and sequencing were performed on a subset of PCR products to confirm that only bla CMY-2 was amplified in samples with the correct melt temperature. The log-scale amplification plots were inspected to confirm that all samples with an amplified product were in the exponential phase of amplification when the threshold was crossed. Any spurious results due to non-specific amplification or well evaporation were excluded prior to threshold calculations.
A single consolidated standard curve was created by ordinary least squares regression using the the MxPro (v.4.10) default method for multiple run projects. The quantities of bla CMY-2 in the unknowns were estimated from the standard curve that ranged from 1×105 copies to 1×101 copies. MxPro (v.4.10) software was used to calculate thresholds, Cq's, the consolidated standard curve, and bla CMY-2 quantities in the unknowns.
Statistical analysis
To determine if ceftiofur treatment had a significant effect on the quantity of bla CMY-2 over time, we utilized a hierarchical model that was previously developed to analyze censored qPCR data (Boyer et al., 2012). The lower limit of quantification (LOQ) of the qPCR assay was determined to be 10 gene copies per 5 μL of DNA solution. When a qPCR well shows no amplification (a value less than the LOQ), many researchers will record the result as 0 gene copies; in reality, a value less than the LOQ may indicate that there is gene in the entire sample, but that the actual concentration is less than the sensitivity and resolution of the assay, and therefore, the result is considered to be censored. The hierarchical model we developed estimates the mean log10 gene quantity in the 0.1-g fecal sample from the observed gene quantities measured in triplicate and determines the most likely estimate of gene copies per gram for each censored observation.
In this model, the unknown log10 gene quantity in sample j (yj ) is assumed to follow a normal distribution with a mean estimated by the regression equation β0k +β1k *day+β2*treatment, where β0k is a random intercept for animal k, β1k is a random slope over time for animal k, and β2 is a coefficient for ceftiofur treatment, a time-varying indicator (Gelman and Hill, 2007; McCoach and Kaniskan, 2010). Because all DNA extraction protocols yield less than 100% of the DNA present in a sample due to incomplete cell lysis and volume losses during the extraction process (Martin-Laurent et al., 2001), a correction factor for DNA extraction efficiency is included in the model. The model parameters were estimated by Markov chain Monte Carlo (MCMC) methods using WinBUGS v1.4 (Lunn et al., 2000). Non-informative normal priors were given to all of the coefficients, and non-informative gamma priors were used for the overall precision (inverse of the variance), the precision of the sample means, and the precision of the animal random effects. The model was run with three chains, each with different initial values, for 5000 iterations with a burn-in of 1000 iterations.
Results
Treatment with ceftiofur was significantly associated with bla CMY-2 quantities during treatment (Fig. 1); there were no apparent differences between the two groups in baseline bla CMY-2 levels or levels post-treatment. The differences in bla CMY-2 levels were only significantly different between the treated and untreated cows on days 3 and 5 during treatment. There was minimal inter-animal variation as indicated by the high degree of similarity in the animal random intercept (data not shown).
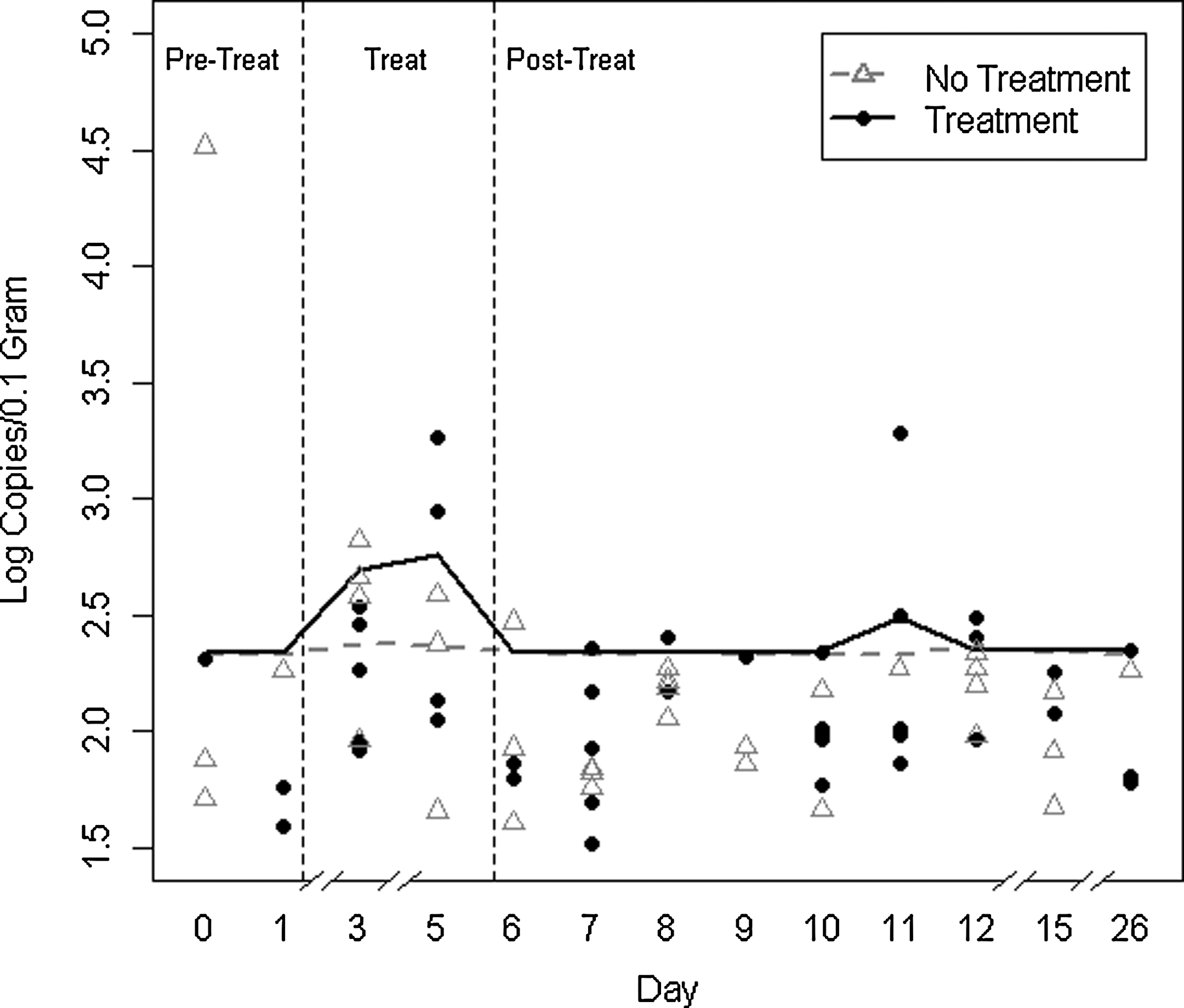
Fitted group means of log10 bla CMY-2 quantity per 0.1 g of sample in cows treated with ceftiofur (solid line) on days 1–5 and untreated cows (dashed line) by day of sample collection, and bla CMY-2 quantities of samples (average of triplicate observations) estimated by the standard curve of treated (circles) and untreated (triangles) animals.
A total of 390 qPCR reactions representing 130 fecal samples were run in six assays. A threshold of 0.15 was used for the consolidated standard curve which had a calculated efficiency of 1.01, R 2 of 0.990, and y-intercept of 35.78. The efficiencies of the six individual standard curves had a mean of 0.9905 (95% confidence interval [CI], 0.9902–0.9908). Among the 390 triplicate reactions that were performed, 46 (11%) were excluded prior to threshold calculations because their dissociation curves showed evidence of primer-dimer formation or some other non-specific product. No single sample had all three of its triplicates excluded for this reason.
Among the remaining 354 reactions, 233 (60%) had no Cq and showed no sign of any amplification. The remaining 122 reactions had dissociation curves that showed a single product with the correct melting temperature within a range of 85.6–86.2°C. The bla CMY-2 copy number per positive qPCR reaction ranged from 41 copies down to one copy. Five reactions had copy numbers estimated by the standard curve to be less than one. Reactions with gene quantity estimated to be less than 10 or with no Cq (84% of the observations) were considered censored and their values were set to missing so that they could be imputed by the statistical model. There was one sample from the untreated group on the first day of the study (day 0) where the quantities of all three triplicates were at least 10-fold higher than all other samples. Including this outlier in the analysis had a large influence on the results; therefore, the results presented here exclude the triplicates from that one sample. Although the small aliquot of feces used from that single animal may have contained a high quantity of bla CMY-2, the inclusion of this sample in the analysis would have resulted in an overall non-significant relationship of treatment status with bla CMY-2 copy number.
Of the 129 fecal samples analyzed, 79 (61%) had at least one of its triplicates test positive for bla CMY-2. At least two cattle from each treatment group were positive for bla CMY-2 on every sampling day of the study except for days 8 and 26, when only one positive sample was detected in the untreated group. Every animal in the study tested positive for bla CMY-2 at least one time, but no animals were positive on every day of the study. Five of 10 samples from treated animals and four of 10 samples from untreated animals were positive on the 2 days prior to treatment.
The crude bla CMY-2 quantities estimated by the standard curve increased in both the ceftiofur-treated and untreated groups during the treatment period and then returned to near pre-treatment levels on the first day post-treatment (Fig. 1). Gene quantities were variable in both groups for the remainder of the post-treatment phase of study. Gene quantities during the post-treatment period were similar to pre-treatment levels in both groups, with the exception of a 1-day spike in bla CMY-2 quantity on day 11 in the treatment group due entirely to a high bla CMY-2 quantity in one cow (Fig. 1).
Discussion
The previous study of this cohort by Singer et al. (2008) presented several pieces of evidence to support the conclusion that bla CMY-2 was present in both treatment groups throughout the course of the study. The authors concluded that the higher frequency of resistant E. coli that was observed in the treated group was likely due to a temporary reduction of a much larger ceftiofur susceptible E. coli population that permitted the detection of resistant E. coli. The authors also showed that the genetic diversity of the E. coli population at the end of the study was the same as it was prior to treatment, suggesting that resistant E. coli did not exploit the temporary competitive advantage offered by the ceftiofur treatment. The results of our analysis showed a statistically significant spike in the quantity of bla CMY-2 during the treatment period, indicating that not only was it temporarily easier to detect bla CMY-2 in cultured E. coli isolates from treated animals but the quantity of the gene may have temporarily increased in the bacterial population as well. It is unknown, however, whether this was due to an increase in the bla CMY-2 positive E. coli population. Regardless, the results suggest that the administration of ceftiofur to these animals resulted in the short-term selection of the bla CMY-2 gene, and that the small increase in the non-treated group was likely due to bacteria being shared among commingled animals.
An interesting difference between our results and the results from the study of bla CMY-2 quantities in feedlot cattle is the higher quantities of bla CMY-2 that were observed in all treated and untreated groups in that study (Alali et al., 2009). The lowest group mean that was reported in their data was nearly 2 log10 higher than the highest group mean that we observed. The data presented here included a high proportion of censored observations, and these results are consistent with what was observed in this cattle population when tested for bla CMY-2 by end-point PCR of community DNA (Singer et al., 2008). It is unknown to what degree reactions below the LOQ affected the results of the feedlot study. In addition, the quantities reported here were estimated assuming 65% DNA extraction efficiency which may be overly optimistic (Mumy and Findlay, 2004).
Another possible explanation for the difference between the two studies may relate to study design. Our study was an observational study with five treated animals mixing freely among a herd of approximately 150 milking cows. The feedlot cattle study was an experimental design of 61 animals with approximately half receiving treatment. The higher density of treated animals in the experimental study may have resulted in more shedding of resistant bacteria that could spread to other cattle in the study population (Hancock et al., 1998). We did notice a small increase in gene quantity in the untreated group, perhaps due to shared bacteria among the commingled animals. In studies of antibiotic resistance transmission, the overall prevalence of resistance as well as the amount of selection pressure being applied among hosts in the population is critical in predicting the degree to which the antibiotic will impact the apparent resistant population. However, it may also be the case that bla CMY-2 quantities are generally higher in feedlot cattle than in dairy cattle due to differences in their respective environments and management methods. The prevalence of phenotypic resistance in E. coli from that population of feedlot cattle was also higher than what was observed in the dairy cattle in this study (Lowrance et al., 2007). We do not believe that differences in the results between these studies were due to the presence of PCR inhibitors in our samples as both this study and the feedlot cattle study extracted DNA using the same commercial kit.
Conclusion
This study demonstrated that treatment of diseased dairy cattle with ceftiofur resulted in a transient increase in the bla CMY-2 gene quantity in the lower intestinal bacterial population of those cattle. Gene quantities returned to pre-treatment levels immediately after treatment suggesting that treatment did not have a lasting effect on the bacterial population. This conclusion is in agreement with a previous study of these cattle that focused on ceftiofur resistance of cultured E. coli (Singer et al., 2008). We also observed a slight increase in bla CMY-2 quantity in untreated cattle which may be indicative of transmission of resistant bacteria among these cattle. Resistant E. coli were not isolated from untreated animals in the previous study suggesting that this cultivation independent approach focusing on community DNA may be a more sensitive method for measuring low levels of antibiotic resistance than one that focuses on particular species of bacteria.
Footnotes
Acknowledgments
We thank Janet Anderson, Kjerstie Wiltzen, and Lauren Petersen for technical and laboratory assistance. This project was supported by the National Research Initiative (grants 2003-35212-13853 and 2004-35212-14885) from the USDA National Institute of Food and Agriculture, Epidemiological Approaches for Food Safety–32.1.
Disclosure Statement
No competing financial interests exist.