
Abstract
Sodium metasilicate (SMS) is a U.S. Department of Agriculture–approved antimicrobial for use in meat and poultry processing and has been known to be effective against various foodborne pathogens. However, its antimicrobial mechanism has not yet been revealed. In this study, we attempted to elucidate the mechanism by which SMS inactivates Listeria monocytogenes, a Gram-positive bacterial pathogen encountered commonly in ready-to-eat meat and poultry products. L. monocytogenes (Scott A) cells were treated with different concentrations of SMS (1.0, 2.0, 3.0, 4.0, 5.0, and 6.0% [wt/vol]) and compared with high pH treatment (0.1, 0.2, and 0.3N NaOH solutions) for 1, 10, and 30 min. SMS exhibited concentration and time effects on inactivation of L. monocytogenes. The effect of SMS on the membrane integrity and viability of L. monocytogenes was determined by use of propidium iodide (PI) and SYTO9 nucleic acid stains with subsequent flow cytometry. The breakage in membrane integrity was observed by uptake of PI by cells treated with SMS with subsequent flow cytometry. Ultrastructural changes from corresponding transmission electron micrographs further revealed the disruption in the cytoplasmic membrane and changes in the morphology of the cells treated with SMS and high pH. The results from flow cytometry experiment and transmission electron microscopy study indicated that following SMS treatment, the membrane integrity of L. monocytogenes was compromised leading to leakage of intracellular contents and subsequent cell death.
Introduction
L
Recent USDA-FSIS data indicate that the prevalence of L. monocytogenes in RTE meat and poultry products has declined, but still the occurrence of L. monocytogenes in RTE meat and poultry products is a significant health hazard. Approximately 13 different serotypes of L. monocytogenes have been described, but the most common serotypes implicated in foodborne illnesses and outbreaks are 1/2a, 1/2b, and 4b (Kathariou, 2002; Gorski et al., 2006). The presence of L. monocytogenes in RTE products usually involves post-processing contamination and is of high significance in terms of food safety. Despite vast research involving use of different antimicrobials such as organic acids and their salts, bacteriocins such as nisin, and natural antimicrobials such as rosemary to restrict and eliminate L. monocytogenes in RTE food products (Burt, 2004; Lungu and Johnson, 2005; Over et al., 2009; Ruiz et al., 2009), this pathogen has been encountered quite frequently; which endangers consumer health, especially the at-risk populations. Therefore the implementation of food safety processes that will control the occurrence of L. monocytogenes in RTE foods continues to be a major concern and need for the food industry. L. monocytogenes has been known to be susceptible to highly alkaline conditions (pH 11.0–12.4) (Mendonca et al., 1994; Taormina and Beuchat, 2002).
Sodium metasilicate (SMS) is a highly alkaline chemical and has been approved for its use as an antimicrobial in RTE meat and poultry products as a 6% solution applied on the surface of the product at levels not exceeding 300 ppm of the finished product (USDA-FSIS, 2012). SMS was effective in reducing Gram-negative foodborne pathogens such as Salmonella and Escherichia coli O157:H7 in vitro and in beef trimmings, beef hide, and chicken breasts (Weber et al., 2004; Carlson et al., 2008; Pohlman et al., 2009; Sharma et al., 2012). However, very little is known about its antimicrobial efficacy against Gram-positive pathogens like L. monocytogenes and also its antimicrobial mechanism of action.
The objectives of the present study were to determine the antimicrobial efficacy of SMS and high pH against L. monocytogenes in vitro and to elucidate the mechanism of antimicrobial action of SMS against L. monocytogenes, if any. The combination of flow cytometry with fluorescent stains has been used to assess variety of cell functions such as cell viability based on membrane integrity and other physiological functions (Davey and Kell, 1996; Gruden et al., 2004; Paparella et al., 2008; Kim et al., 2009). In the present study, we used propidium iodide (PI) and SYTO9 dyes with flow cytometry and transmission electron microscopy (TEM) to assess the effect of SMS and high pH on changes in membrane integrity and intracellular structures of L. monocytogenes.
Methods
Bacterial culture and exposure of cells to SMS and high pH
Reference strain of L. monocytogenes (Scott A) obtained from ABC Research Corporation (Gainesville, FL) was used in this study to evaluate the antimicrobial properties of SMS. Stock cultures were prepared by transferring the reference strain to test tubes containing 10 mL of tryptic soy broth (TSB), followed by incubation at 35°C for 24 h. After incubation, the cultures were centrifuged in sterile 15-mL centrifuge tubes (model 05-539-5; Fisher Scientific, Pittsburgh, PA) at 5,000 rpm for 10 min at 4°C (Sorvall RC-5; Dupont Instruments, Newton, CT). The supernatant was discarded, and the pellets were resuspended in 10 mL of sterile 0.1% peptone water (CM009; Oxoid Ltd., Basingstoke, UK), centrifuged again, and pellets were suspended in 1 mL of TSB supplemented with 30% glycerol in a 2-mL cryogenic vial (model 430488; Corning Inc., Corning, NY), stored at −80°C, and used as the stock culture for the inoculation studies. The working cultures were prepared by thawing the deep-frozen culture at room temperature for 5–10 min, and a loopful was streaked onto tryptic soy agar (TSA) (MP Biomedicals, LLC, Solon, OH), followed by incubation at 35°C for 24 h. One colony from the working culture was selected and transferred to 10 mL of TSB and incubated at 35°C for 20 h. Cells were pelleted by centrifugation at 5,000 rpm for 10 min at 4°C, washed, centrifuged again, and re-suspended in 10 mL of treatment solution at room temperature containing either (i) 0.1% sterile peptone water (control); (ii) 1.0%, 2.0%, 3.0%, 4.0%, 5.0%, and 6.0% (wt/vol) SMS solutions; or (iii) 0.1, 0.2, and 0.3N NaOH solutions for 1, 10, and 30 min.
Cell viability
Treated cells were tested for their viability by taking out 1-mL aliquot from each treated cell suspension and serially diluting in 9 mL of sterile 0.1% peptone water. A volume of 0.1 mL from each dilution was pipetted onto duplicate prepoured modified Oxford medium (MOX) plates prepared by mixing Oxford medium (222530; Becton, Dickinson and Company, Sparks, MD) with modified oxford antimicrobic supplement (211763; Becton, Dickinson and Company) for L. monocytogenes recovery. The MOX agar plates were incubated aerobically for 48 h at 35°C, and after incubation, colony-forming unit (CFU) from each plate were counted, averaged, and reported as log CFU/mL of the sample. Duplicate samples for each treatment were analyzed, and the experiment was repeated three times.
Flow cytometry analysis for the membrane integrity
Fluorescent dyes PI (Invitrogen, Eugene, OR) and SYTO9 (Invitrogen) were used in this study to differentiate live and dead bacterial cells based on the membrane integrity. Both of these dyes bind and stain the nucleic acids. PI is a red fluorescent stain that can only enter and stain the cells presumed to be dead with damaged or permeabilized cytoplasmic membranes. PI is normally excluded by live cells, whereas SYTO 9, which emits green fluorescence, is able to stain both live and dead cells (Kim et al., 2009). After exposure of cells to 1.0%, 2.0%, 3.0%, and 4.0% SMS and 0.1N NaOH for 1 and 30 min, each sample (108 CFU/mL) was serially diluted to 106 CFU/mL in phosphate-buffered saline (PBS; NaCl 8 g/L, KCl 0.2 g/L, Na2HPO4 1.44 g/L, KH2PO4 0.24 g/L; pH 7.4). One milliliter of the diluted sample was then stained with 2.5 μL of PI (1.5 mM) and SYTO9 (0.5 mM) either single or in combination and incubated at room temperature for 30 min. Samples were run in FACSort machine (BD Biosciences) for the flow cytometry analysis with emission wavelengths for SYTO9 and PI at 530±15 nm and 585±21 nm, respectively. Data were analyzed by Cell Quest Software, Version 3.3 (BD Biosciences). The flow cytometry analysis was repeated twice for each treatment, and the results were found to be reproducible. All the samples were analyzed in the flow cytometry section of the Interdisciplinary Center for Biotechnological Research (ICBR; University of Florida, Gainesville, FL).
Transmission electron microscopy
TEM was performed to observe the changes at cellular level following exposure of L. monocytogenes cell suspensions to 2% SMS (for 1 and 30 min) and 0.1N NaOH (for 30 min) treatments. Treated and untreated (control) cells were centrifuged to pellet by centrifugation at 12,000 rpm for 10 min, and washed and fixed in 4% paraformaldehyde + 2% glutaraldehyde in 0.1M sodium cacodylate, pH 7.24. Fixed cells were processed with the aid of a Pelco BioWave laboratory microwave. The samples were washed in 0.1M sodium cacodylate buffer, and encapsulated in molten 3% low-temperature gelling agarose Type IV (A3643; Sigma-Aldrich, Saint-Louis, MO) post fixed with 1% osmium tetroxide (OsO4), water washed, and dehydrated in a graded ethanol series followed by 100% acetone. Dehydrated samples were infiltrated in graded acetone/Spurrs epoxy resin and cured at 60°C. Cured resin blocks were trimmed, thin sectioned, and collected on formvar copper 200 mesh grids, post-stained with 2% aq. Uranyl acetate and Reynold's lead citrate. Sections were examined with a Hitachi H-7000 TEM (Hitachi High Technologies America, Inc., Schaumburg, IL) and digital images acquired with Veleta camera and iTEM software (Veleta-Olympus Soft-Imaging Solutions Corp., Lakewood, CO). All the samples were processed and analyzed in the Electron Microscopy and Bioimaging section of ICBR.
Statistical analysis
Data for the cell viability experiment were analyzed by the analysis of variance (ANOVA) procedure of SAS (SAS Institute, 2009) to determine differences among treatments. The mean separation was performed using Tukey's method, and a level of significance of α=0.05 was used to determine any significant differences among means.
Results and Discussion
Effect of SMS and high pH on cell viability
This study revealed that SMS had concentration and time effect on inactivation of L. monocytogenes in suspension (Table 1). No significant reduction in L. monocytogenes (p>0.05) was observed with 1-min exposure to 1% SMS. Concentrations of 1%, 2%, and 3% SMS reduced L. monocytogenes by more than 5 log after 30 min. Exposure for 1 min to 4% and 5% SMS reduced L. monocytogenes by more than 5 logs. L. monocytogenes was not detected after 1-min exposure to 6% SMS, after 10-min exposure to 5% and 6% SMS, and after 30-min exposure to 4%, 5%, and 6% SMS. Except for 1-min exposure of L. monocytogenes to 4% SMS, treating L. monocytogenes with 0.1, 0.2, and 0.3N NaOH for 1, 10, and 30 min resulted in similar reductions in L. monocytogenes (Table 2). One-minute exposure of L monocytogenes to 3% (pH 12.8) and 4% (pH 12.9) SMS resulted in 3.04 and 5.16 log reductions, respectively, while 1-min exposure of L. monocytogenes to 0.2N (pH 12.8) and 0.3N (pH 12.9) NaOH resulted in 1.23 and 1.96 log reductions, respectively. As exposure time of L. monocytogenes to 4% SMS and corresponding 0.3N NaOH treatments increased to 30 min, L. monocytogenes was not detected. The data revealed that high pH exerted a concentration and time effect on reduction of L. monocytogenes in vitro. In a similar study, exposure of L. monocytogenes to high pH (12.0) for 15 min resulted in only 1 log unit reduction (Mendonca et al., 1994). In an earlier study, we found that SMS was unable to exert any antimicrobial effects against Salmonella following pH neutralization (data not shown) of 1% and 2% SMS solutions, which indicates that it could be the high pH of SMS that is responsible for bactericidal effects against Salmonella Typhimurium. As evident from findings in this study when high pH solutions of NaOH were used corresponding to pH values of 2%, 3% and 4% SMS solutions, similar responses were observed in terms of reduction of L. monocytogenes cell numbers in suspension (Table 2).
Means within a column lacking a common superscript differ (p<0.05).
n=6 values per mean.
CFU, colony-forming unit; SMS, sodium metasilicate; ND, not detected.
Means within a column lacking a common superscript differ (p<0.05).
n=6 values per mean.
CFU, colony-forming unit; ND, not detected.
Flow cytometry analysis of SMS and high pH–treated cells
The effect of SMS (different concentrations and time of exposure) on the membrane integrity of L. monocytogenes was determined by use of PI and SYTO9 with subsequent flow cytometry. The fluorescent signals obtained from the PI and SYTO9 after treatment of L. monocytogenes with different SMS concentrations for 1 and 30 min are presented in two-dimensional dot plots (Figs. 1 and 2). The cell population on the dot plot was clustered and identified in three different zones that include PI positive, SYTO9 positive, and unstained. The cell populations present in the PI-positive zone were stained with PI, and their cytoplasmic membrane integrity was compromised, suggesting that this population of L. monocytogenes was dead. It should be noted that as the concentration of SMS increased from 2% to 4% at exposure time of 1 min, there was an increase in PI-positive cell population for a corresponding increase in SMS concentrations (Fig. 3A). These findings corroborate the observations from plate counts, where we found that increasing the SMS concentration resulted in more inactivation of L. monocytogenes cell populations. It was also observed that the intensity of fluorescent signals of PI-positive cells increased when PI and SYTO9 were added together as a mixture as compared to PI added alone for each treatment (Fig. 3A). It probably could be due to the quenching effect or the transfer of fluorescence energy between SYTO9 and PI dyes when they were added together (Stocks, 2004; Berney et al., 2007). When used alone, SYTO9 did not provide any significant information on cells with compromised membranes or live cell populations. However, when L. monocytogenes cells were exposed to different concentrations of SMS for 30 min, there was a decrease in PI-positive cell populations for a corresponding increase in SMS concentrations (Fig. 3B). The possible explanation for this effect could be the longer exposure of L. monocytogenes to harsh alkaline conditions generated by elevated SMS concentrations resulting in more damage to cytoplasmic membrane and leakage of cytoplasmic contents (including nucleic acids), thereby producing a low-intensity signal from red fluorescent PI dye. PI binds with double-stranded nucleic acids with a stoichiometry of one dye molecule per four to five base pairs of DNA in order to produce red fluorescent signals. Hence, more damage and loss of DNA following exposure to SMS for longer duration might not have allowed PI to stain enough DNA to cause high-intensity red fluorescent signals.
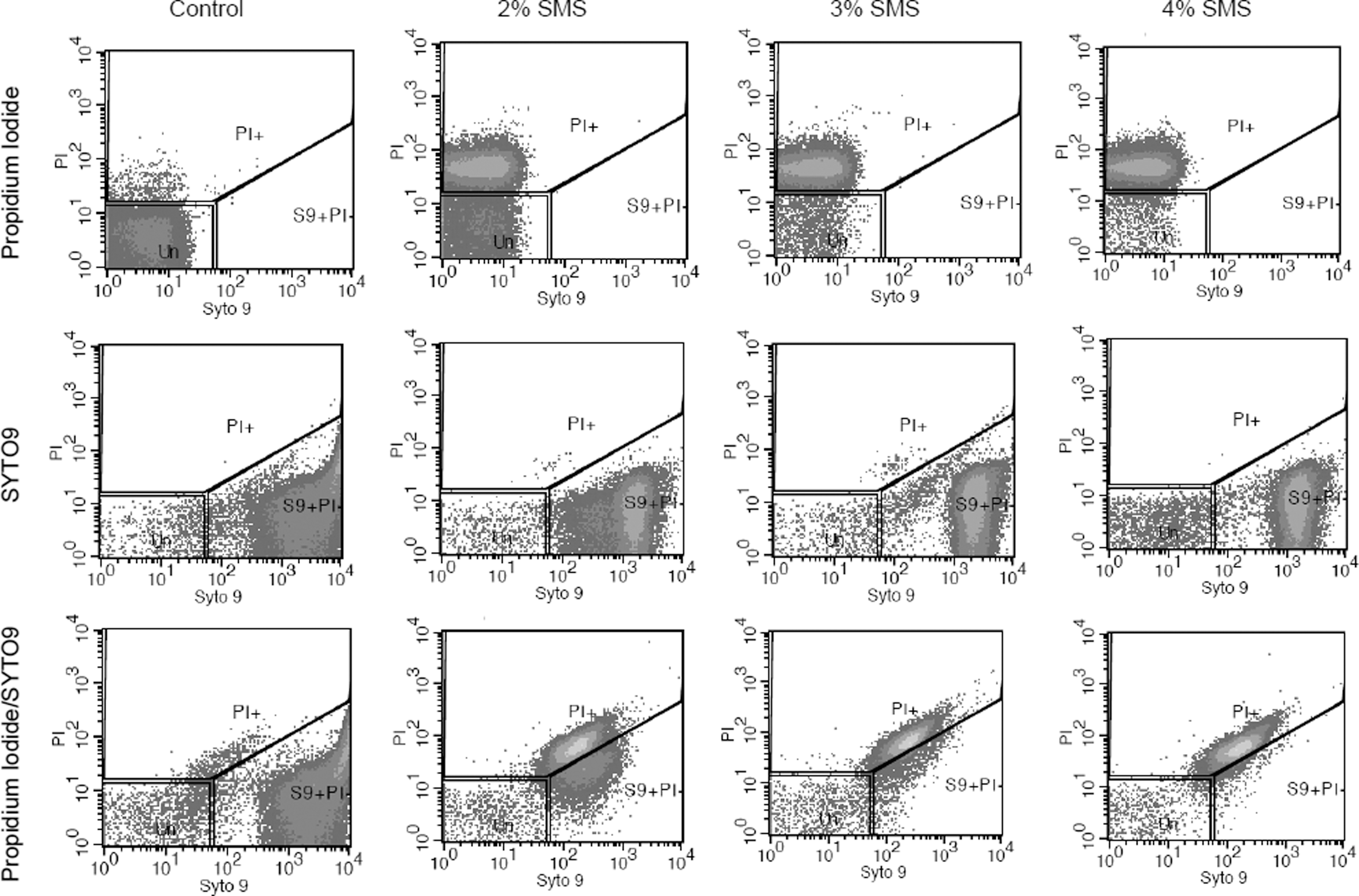
Flow cytometry analysis of Listeria monocytogenes (Scott A) treated with sodium metasilicate (SMS) and stained with PI, SYTO9, and PI/SYTO9. L. monocytogenes cells were untreated (control) or treated with SMS at the indicated concentrations and time exposure for 1 min at room temperature. The blocked regions PI+, S9+PI-, and Un represents the PI-positive cells (loss of membrane integrity), SYTO9-positive cells, and unstained cells, respectively.
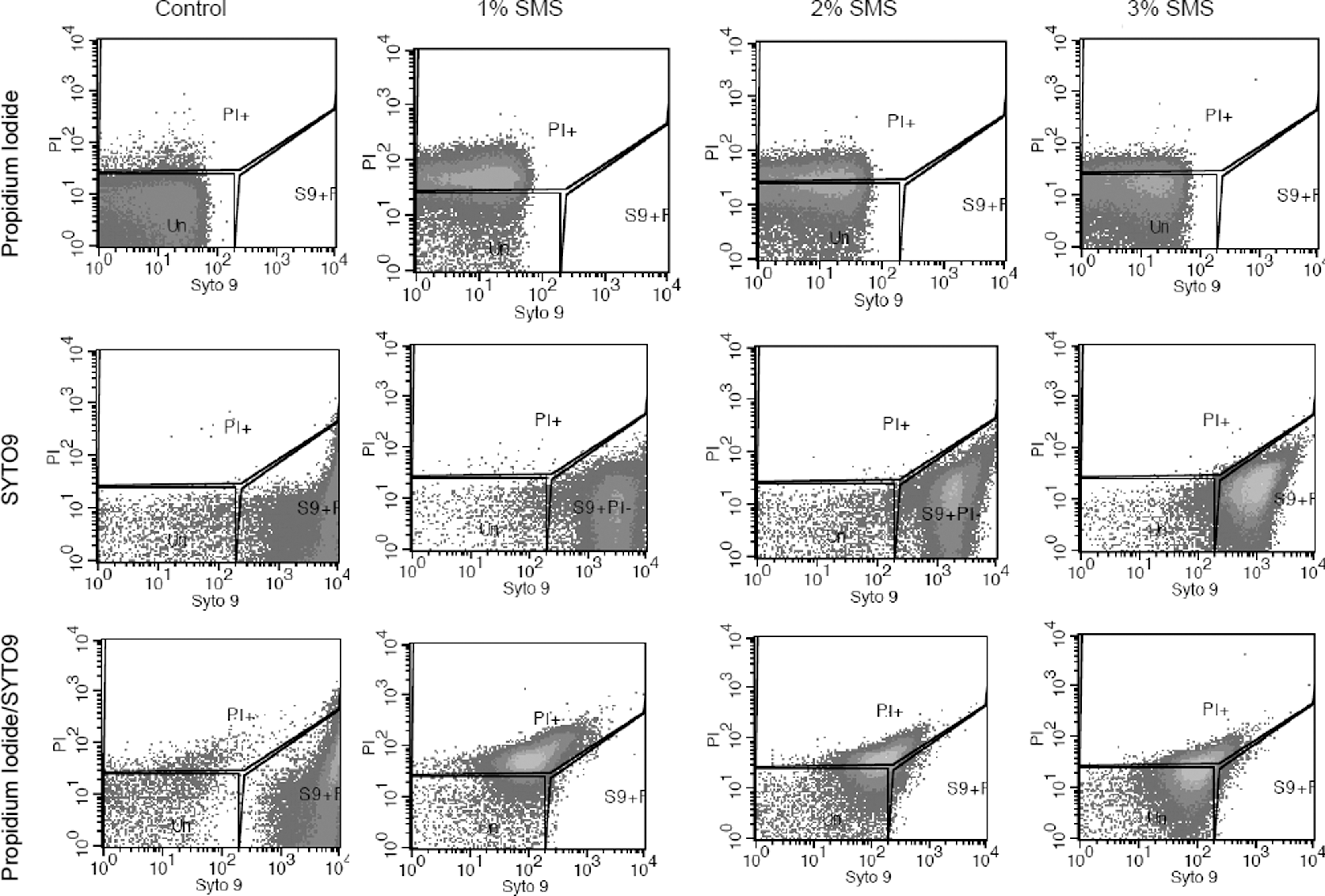
Flow cytometry analysis of Listeria monocytogenes (Scott A) treated with sodium metasilicate (SMS) and stained with PI, SYTO9, and PI/SYTO9. L. monocytogenes cells were untreated (control) or treated with SMS at the indicated concentrations and a time exposure for 30 min. The blocked regions PI+, S9+PI-, and Un represents the PI-positive cells (loss of membrane integrity), SYTO9-positive cells, and unstained cells, respectively.
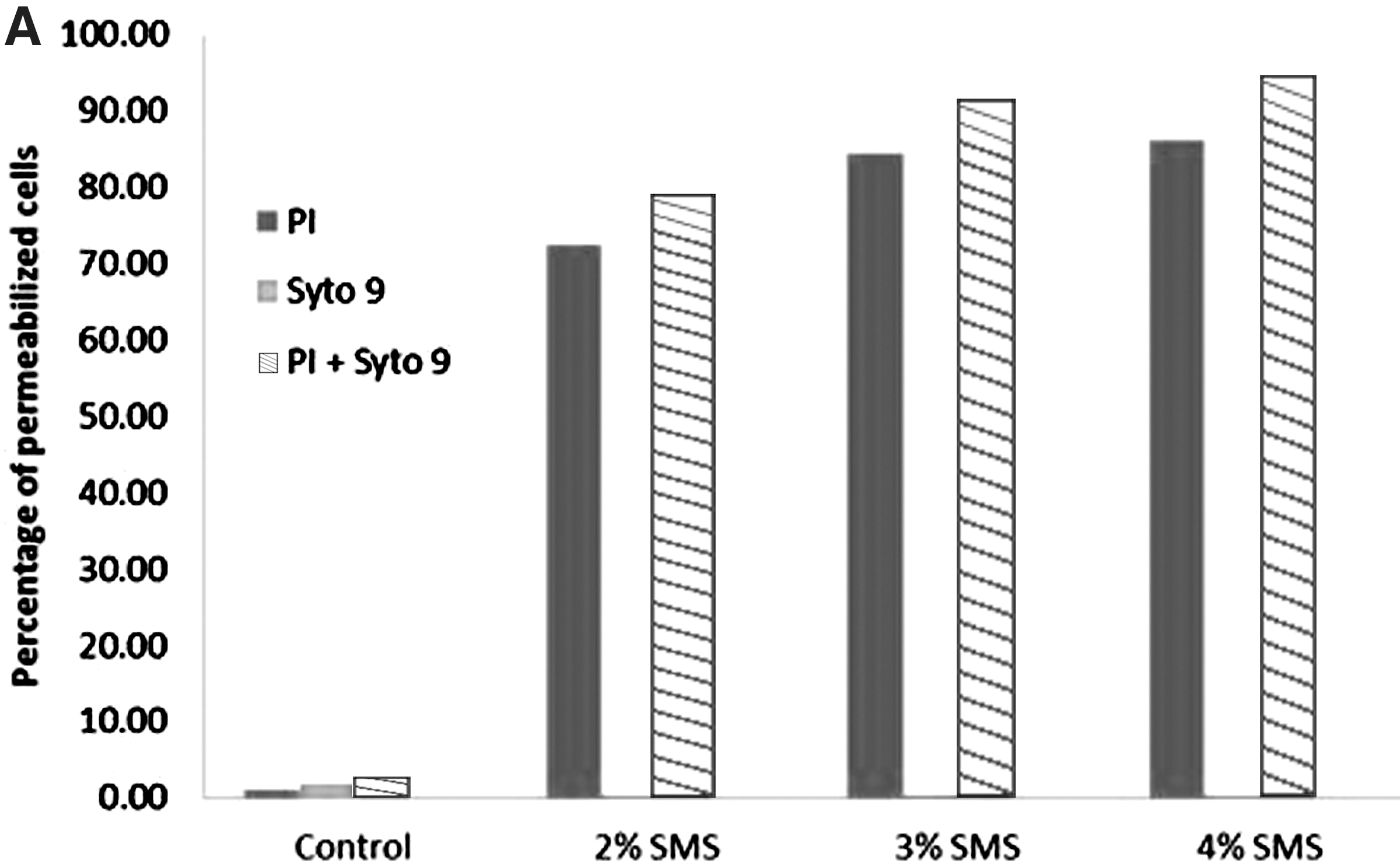
The percentage of membrane permeabilized Listeria monocytogenes cells (PI positive) following treatment with different concentration of sodium metasilicate (SMS) at room temperature for 1 min
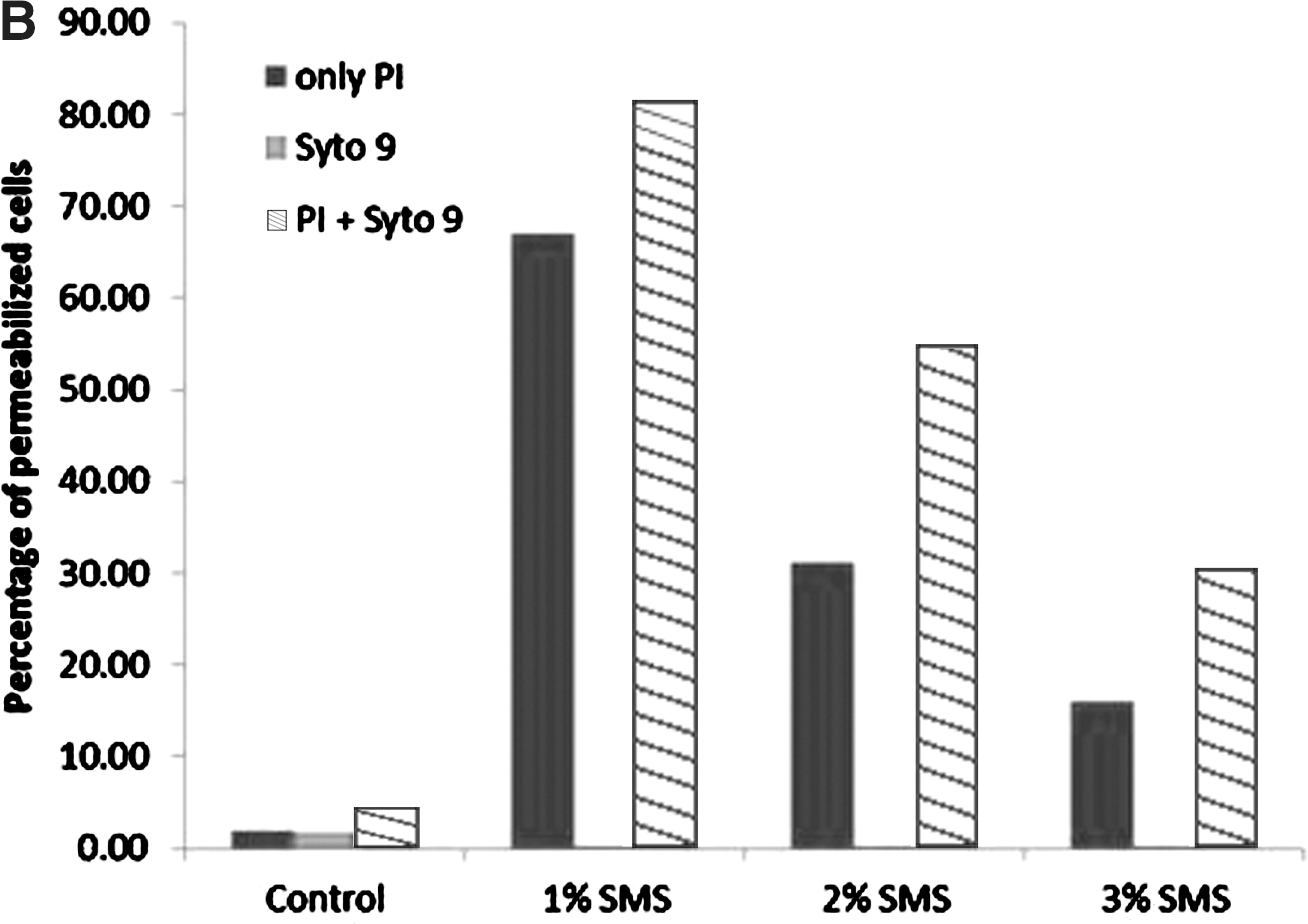
The results from flow cytometry experiment, particularly the 1-min exposure of L. monocytogenes to SMS, matched with those of the plate count experiment, thereby confirming that there was a concentration effect of SMS on viability of L. monocytogenes. However, data from flow cytometry experiment under which L. monocytogenes cell populations were treated with different concentrations of SMS for 30 min did not match the observations from plate count experiments. This observation indicates that flow cytometry did not provide reliable information on differentiation of live and dead L. monocytogenes cells by using PI/SYTO9 combination particularly when exposed to different concentrations of SMS for longer duration. It has been reported in the literature that combining SYTO9 and PI does not always provide distinct differentiation of live and dead bacterial cell populations with much accuracy (Berney et al., 2007; Kim et al., 2009). Similar findings were observed for flow cytomerty analysis for L. monocytogenes treated with high pH (data not shown). In this study, we report the use of PI and SYTO9 and flow cytometry to determine the efficacy of SMS on membrane integrity and differentiation of live and dead L. monocytogenes cell populations.
Transmission electron microscopy
Although L. monocytogenes (Scott A) cells that were exposed to 2% SMS and 0.1N NaOH had different intracellular morphology than untreated cells, the treated cells retained their overall shape (Fig. 4). Cells treated with SMS lost the normal electron dense cytoplasm, which became lighter after the treatment. In addition, cell wall and cytoplasmic membrane of some cells appeared to be disrupted particularly following 30-min exposure to SMS and NaOH as compared to the untreated cells (Fig. 4). There were distinct clear and dark zones seen in the cytoplasm of L. monocytogenes cells after treatment with 2% SMS for 1 min and for 30-min exposure to NaOH. However, for L. monocytogenes treatment with 2% SMS for 30 min, minimal clear and dark zones were observed. In contrast, the cytoplasm of untreated cells was homogenous with thick intact cell walls and indistinct cytoplasmic membranes. Cells exposed to 0.1 N NaOH (pH 12.5) corresponding to pH of 2% SMS solution showed some dissimilarities as compared to cells treated with 2% SMS. In a similar study, after exposure of L. monocytogenes cells to an alkaline environment (pH 12.0), the cytoplasm had distinct clear and dark zones, with bulging of cytoplasmic membrane against the cell wall but no discharge of intracellular material (Mendonca et al., 1994). In contrast to this study, previously we found that SMS caused extensive lysis of Gram-negative Salmonella Typhimurium cells. There was widespread leakage of cytoplasmic contents as observed on transmission electron micrographs. The effect of SMS and high pH on L. monocytogenes, which is Gram positive, is not as extensive as seen with Salmonella in the previous study. This difference in findings indicates that Gram-negative and Gram-positive bacteria most likely differ in their sensitivities to SMS owing to their differences in their cell wall structures (Mendonca et al., 1994). As reported earlier by Mendonca et al. (1994), the thick peptidoglycan layer in the cell walls of Gram-positive bacteria, in this case L. monocytogenes, helps the intact cells to stabilize their weakened cytoplasmic membrane as a result of a highly alkaline environment.

Transmission electron micrographs of Listeria monocytogenes (Scott A) exposed to untreated (control;
In summary, the results from flow cytometry experiment and TEM study indicated that following 2% SMS treatment, the membrane integrity of L. monocytogenes is compromised leading to leakage of intracellular contents and eventually cell death. The breakage in membrane integrity was also seen by uptake of PI by cells treated with SMS and subsequent flow cytometry. In addition, it could be possible that SMS and NaOH act differently in their killing/inactivation mechanism of L. monocytogenes as seen in the differences in electron micrographic findings between SMS and sodium hydroxide treated cells. Further studies may be necessary to reveal the difference in their mechanism of action.
Footnotes
Acknowledgments
We are thankful to the Midwest Poultry Consortium for providing the financial assistance to conduct this study. We also acknowledge the Interdisciplinary Center for Biotechnology Research (ICBR), University of Florida for providing assistance in processing and analyzing samples for flow cytometry and transmission electron microscopy. This article was approved for publication as Journal Article No. J-12138 of the Mississippi Agricultural and Forestry Experiment Station, Mississippi State University. Use of trade names in this publication does not imply endorsement by Mississippi Agricultural and Forestry Experiment Station of these products, nor similar ones not mentioned.
Disclosure Statement
No competing financial interests exist.