
Abstract
Development of antibiotic resistance may alter the virulence properties of bacterial organisms. In this study, nine clinical ceftriaxone-susceptible Salmonella enterica serotype Typhimurium strains were subjected to stepwise selection with increasing concentrations of ceftriaxone in culture media. Mutations in virulence-associated genes and antibiotic efflux genes were analyzed by polymerase chain reaction (PCR) and DNA sequencing. The expression levels of virulence genes invA and stn as well as efflux pump genes tolC, arcA, and arcB before and after the selection were measured by real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR). The stepwise selection resulted in the development of Salmonella strains that were highly resistant to ceftriaxone. Sequence analysis did not reveal any mutations or deletions in the examined virulence genes and regulatory gene, but a silent mutation (T423C) in acrR (encoding a repressor for the efflux pump) was detected in most of the ceftriaxone-resistant strains. The qRT-PCR revealed increased expression of the AcrAB-TolC efflux pump and decreased expression of invA and stn in the ceftriaxone-resistant strains. Moreover, decreased invasion into cultured epithelial cells and reduced growth rates were observed with the resistant strains. These results suggest that acquisition of ceftriaxone resistance is associated with the overexpression of the AcrAB-TolC efflux pump and leads to reduced virulence in Salmonella Typhimurium.
Introduction
In addition, the expression of genes required for invasion is tightly regulated by a variety of regulatory mechanisms and environmental conditions such as temperature, pH, and antimicrobials (Jones, 2005; Rhen and Dorman, 2005). Previous studies have shown that there is a possible link between fluoroquinolone resistance and decreased cell invasion (Fabrega et al., 2009; O'Regan et al., 2010). It was hypothesized that the resistance–conferring mutations in the DNA gyrase gene (gyrA) reduce DNA superhelicity, altering the expression of genes involved in invasion (Fabrega et al., 2009; Galan and Curtiss, 1990; Wang et al., 2009).
The third generation cephalosporins (ceftriaxone or cefixime) are one of the most effective treatments for invasive diseases (Cherubin et al., 1986). However, emergence and dissemination of β-lactam resistance genes have compromised treatment, which has become a serious problem for public health worldwide (Pfeifer et al., 2010). The main mechanisms of cephalosporin resistance in Salmonella have been attributed to extended-spectrum β-lactamases (ESBLs) and increased expression of efflux pumps, especially the AcrAB-TolC efflux system (Li and Nikaido, 2009).
The AcrAB-TolC tripartite efflux system belongs to the resistance-nodulation-division (RND) family of transporters and has been shown to export a broad range of substrates across both the inner and outer membrane to the extracellular space (Baucheron et al., 2004). Moreover, the AcrAB-TolC efflux system also plays an important part in the invasion of the intestine, because of its involvement in export of host-derived substrates such as bile salts, fatty acids, and steroid hormones, thereby enabling bacteria to survive within the hostile environment of the host (Blair and Piddock, 2009; Buckley et al., 2006; Virlogeux-Payant et al., 2008). The tolC mutant of Salmonella Typhimurium poorly adhered to both human embryonic intestine cells and mouse monocyte macrophages in vitro and was unable to invade macrophages (Buckley et al., 2006; Virlogeux-Payant et al., 2008).
The effect of acquiring cephalosporin resistance on the virulence properties of Salmonella is still poorly defined. In this study, we hypothesize that development of cephalosporin resistance may be attributed to the up-regulation of the AcrAB-TolC efflux pump system. Additionally, we hypothesize that stepwise antibiotic selection may affect the general physiological state of Salmonella, leading to reduced virulence. Thus, the main objective of this study was to investigate the relationship between expression of the AcrAB-TolC efflux pump and the enhanced resistance to the third-generation cephalosporins, as well as the association between the expression of virulence genes invA and stn and the invasiveness of ceftriaxone-resistant Salmonella Typhimurium strains.
Materials and Methods
Bacterial strains and selection of resistant mutants
Nine cephalosporin-susceptible Salmonella Typhimurium strains used in this study were isolated from stool samples of diseased ducks in Guangdong province in China. All isolates were biochemically characterized by the API 20E system (BioMerieux, Marcy-l'Etoile, France) and serotyped by slide agglutination test using Salmonella antisera (China Institute of Veterinary Drug Control, Beijing, China). Screening for the presence of ESBLs in Salmonella Typhimurium was performed according to the Clinical and Laboratory Standards Institute (CLSI, 2008) guidelines. All isolates were used as the parental strains. Cephalosporins-resistant strains were obtained in a multi-step selection process in the presence of ceftriaxone. Strains were grown at 37°C on Muller-Hinton plates. Ceftriaxone was only used during the selection processes, starting at half of the minimal inhibitory concentration (MIC) and increasing twofold each step, until reaching a maximum concentration of 64 μg/mL. Colonies were selected to be grown at the consecutive ceftriaxone concentration at each step, and samples were stored simultaneously. The nine parental strains were subjected to the same incubation process in the absence of ceftriaxone as negative controls in parallel.
Antimicrobial susceptibility testing
MICs for ampicillin, ceftriaxone, and ceftazidime were determined by the agar dilution method following the guidelines of CLSI. The MICs were evaluated by using the breakpoints of the CLSI. Escherichia coli ATCC 25922 was used as a quality control organism.
Design of primers for quantitative reverse transcription–polymerase chain reaction (qRT-PCR) and gene sequencing
The qRT-PCR of virulence genes invA and stn, of the efflux pump genes tolC, arcA, and acrB, and of the internal control gene 16S rRNA was performed for all Salmonella Typhimurium strains (parent, resistant mutants, and negative controls) using primers as shown in Table 1). Sequencing of the invasion regulatory genes (hilA, hilC, and hilD) (Jones, 2005) and the AcrAB-TolC transcriptional regulators (soxRS, marRAB, acrR, and ramR) (Abouzeed et al., 2008; Olliver et al., 2004) was accomplished using primers as shown in Table 1. Primers used for the other virulence genes (spvB, spvC, spvD, sopE, phoP/O, sodCI, sodCII, iroB, agfA, hin/H2, repFIIA, and slyA) and ESBLs-encoding genes (including bla
TEM, bla
SHV, bla
DHA and bla
CTX-M) are described elsewhere (Brinas et al., 2003; del Cerro et al., 2003; Guillard et al., 2011; Karasova et al., 2009; Ma et al., 2009; Park et al., 2006; Robicsek et al., 2006). The target genes were amplified by PCR, the products were directly sequenced, and the sequences were compared with those in the GenBank nucleotide database using the BLAST program, available from the National Center for Biotechnology Information (
Growth curve
The growth of a randomly selected strain before and after antibiotic selection was compared. It was monitored by measuring optical density at 600 nm (OD600) in Luria-Bertani (LB) broth. Briefly, the bacteria were initially cultured on LB agar at 37°C overnight. Then the colony was selected and inoculated into 4 mL of LB broth. A 1-mL aliquot of LB broth of overnight culture was inoculated into 100 mL of LB broth without ceftriaxone and cultured at 37°C with rotary shaking at 200 rpm. The OD600 was measured every hour for 17 h. Three independent assays were performed for the strains.
Total RNA extraction and reverse transcription
All strains were incubated in LB broth without ceftriaxone at 37°C overnight. The RNA was extracted from cultures with OD600 at 0.5. Total bacterial RNA was obtained using RNAiso Plus agent and possible DNA contamination was removed by treating with DNase I (RNase-free) according to the manufacturers' instructions. The purity and concentration of RNA was determined by measuring the OD using BioPhotometer plus (Eppendorf, Shanghai, China). A general PCR using the RNA as a template was performed to determine residual DNA. The total RNA was diluted to a final concentration of 1 μg mL−1 prior to reverse transcription. The cDNA was synthesized in a 50-μL reaction mix containing 1 μg of RNA, 10 μL of 5×reverse transcriptase buffer, 50 mM of each dNTP, 50 U of RNase inhibitor, 0.25 μM of Oligo (dT)18 primer, and 50 U of reverse transcriptase M-MLV (Rnase H −). All agents were supplied by TaKaRa (TaKaRa Bio, Dalian, China).
Real-time RT-PCR
The qRT-PCR was performed with SYBR® Premix Ex Taq™ (TaKaRa Bio) in a thermal cycler (iQ5; Bio-Rad, Hercules, CA) according to the manufacturer's instructions. The condition was as follows: denaturing at 94°C for 5 min, followed by 35 cycles at 94°C for 1 min, at 55°C for 1 min, and at 72°C for 1 min, with a final extension at 72°C for 5 min; the melt curve was read from 60°C to 95°C every 1°C at the end. Results were obtained according to the 2-ΔΔCT method, where CT equaled the threshold cycle (Livak and Schmittgen, 2001). All experiments were performed in triplicate, and the relative expression rate was the mean value of nine strains.
Adherence and invasion survival assays
The epithelial cell adherence and invasion assays were performed as previously described (Jouve et al., 1997), with all strains collected before and after antibiotic selection, including negative controls. Briefly, Hela cells in Dulbecco's Modified Eagle's Medium (DMEM; HyClone, Logan, UT) with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY) were cultured onto 24-well tissue culture plates (Corning Inc., Corning, NY) at 1×105 cells per well for 24 h. Hela cells were prewashed three times with Hank's balanced salt solution (HBSS; HyClone) and 1 mL fresh media with 10% FBS was added prior to adherence/invasion assays. Fresh overnight bacteria were washed and diluted in DMEM, and then 100 μL of bacteria was added to each plate at a concentration of approximately 5×107 colony forming units (CFU)/mL. Plates were incubated for 2 h at 37°C with 5% CO2. The monolayers were washed three times with HBSS before disrupting with 1% (v/v) Triton (Sigma, St. Louis, MO). CFUs were determined by serial dilution and plating onto LB agar. For the invasion assay, after allowing the bacteria to adhere to the monolayer, wells were washed three times with HBSS before adding DMEM containing 100 μg/mL gentamicin to kill all external bacteria. Plates were incubated for 2 h at 37°C (5% CO2) and washed three times. One percent (v/v) Triton X-100 was added to the monolayers, and CFUs were determined as described above. Adherence/invasion assays were performed on three separate experiments, with three replications per assay used for each experiment, and the percentage of the number of adhesion/invasive bacteria was reported as the mean value of nine strains.
Statistical analysis
Differences in bacterial growth (log OD600*1000) and epithelial cell adherence/invasion were analyzed with Student's t-test. A p-value of <0.05 was considered statistically significant. An effect on gene expression was considered to be significant when the corresponding ratios were >2.5 or <0.4 (Dumas et al., 2006).
Results
Antimicrobial susceptibility and growth curve
Nine non-ESBLs-producing Salmonella Typhimurium clinical isolates and laboratory-passaged strains were all susceptible to ampicillin, ceftriaxone, and ceftazidime. After stepwise selection with ceftriaxone, the strains showed resistance to ceftriaxone (MIC≥64 μg/mL) and to other β-lactam antimicrobials, including ampicillin and ceftazidime (Table 2). Moreover, under the same culture conditions, the ceftriaxone-resistant strain demonstrated a lower growth rate than its parent strain (p<0.05; Fig. 1).
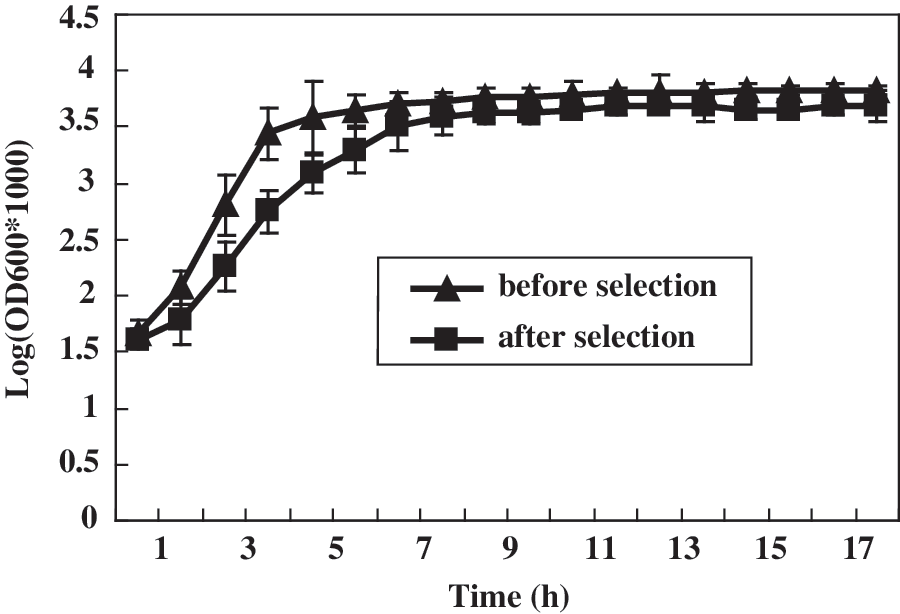
Bacterial growth curves of strain S122 before and after ceftriaxone selection are compared. Measures were taken every hour for 17 h from three independent data. The results are expressed in a semilogarithmic plot. Error bars represent the standard deviation.
CRO, ceftriaxone; AMP, ampicillin; CAZ, ceftazidime.
Detection of ESBLs-encoding genes, virulence, and efflux pump regulatory genes
No ESBLs-encoding genes were detected before or after antibiotic selection. No gene deletions or point mutations were detected in either the target or control strains within the region of invA, stn, and other examined virulence genes. In addition, sequencing results revealed that no point mutations were detected in the invasion regulatory genes hilA, hilC, and hilD. However, a single silent mutation was discovered in the efflux pump regulator acrR, which exerts a repressive effect on AcrAB-TolC expression. A previously undescribed mutation T423C was found in the acrR gene (GenBank accession number JQ689080) in five of the nine resistant Salmonella Typhimurium strains. In contrast, no mutations were detected in the negative controls (strains cultured without antibiotic selection).
Expression levels of virulence genes invA and stn, as well as the AcrAB-TolC efflux pump genes
Expression data revealed that compared with the wild-type ceftriaxone-susceptible strains prior to the stepwise selection, the expression levels of virulence genes invA and stn were significantly decreased (by 24.3- and 9.1-fold, respectively) in the ceftriaxone-resistant strains (Table 3). In contrast, the relative mRNA expression of multidrug efflux pump genes tolC, arcA, and arcB increased 5.4-, 4.0-, and 6.8-fold, respectively, after selection with ceftriaxone. The result indicated that development of resistance to β-lactam antimicrobials was attributable to the up-regulation of the AcrAB-TolC efflux pump in ceftriaxone-resistant strains. No significant changes in the target gene expression were observed in the negative controls (Table 3).
Mean values of nine strains (and the standard deviation) were less than 15% of the mean.
Values represent fold change relative to the transcription level in strains before antibiotic selection.
Adherence and invasion assays
The adhesion and invasion were expressed as a percentage of the number of adhesion/invasive bacteria with respect to the total number of bacteria present in the initial inoculum. The percentage of adherence and invasion significantly decreased from 37.8% and 0.87% for strains prior to antibiotic selection to 5.06% and 0.02%, after selection with ceftriaxone (p<0.01), respectively. Although adhesion of the strains passaged without ceftriaxone also decreased by 2.2-fold (p<0.05), the adhesion of ceftriaxone-resistant strains was much lower than that of negative controls (p<0.01; Table 4).
Mean values of nine strains.
SD, standard deviation.
Discussion
Salmonella, especially serovar Typhimurium, is one of the most important pathogens responsible for gastrointestinal infections in humans and animals (wild and domestic). The third-generation cephalosporins and quinolones are the choice of antibiotics for treating Salmonella infections when MICs are unknown. Fluoroquinolone resistance may appear concomitantly with a loss or decrease in expression of virulence factors and invasiveness in Salmonella Typhimurium (Fabrega et al., 2009). Considering the clinical scenario in which prevalence of ceftriaxone-resistant Salmonella remains low and the AcrAB-TolC efflux pump system plays a key role in non-ESBLs Salmonella Typhimurium (Crump et al., 2011), we hypothesized that acquisition of cephalosporins resistance might repress the expression of virulence genes and decrease the invasiveness of Salmonella.
In this study, we found that the expression levels of two invasion-related genes, invA and stn, were significantly decreased by 24.3- and 9.1-fold, respectively, after stepwise selection with ceftriaxone, whereas no genetic alteration (deletions or mutations) in the invasion-regulating genes were detected, including the main regulator hilA. Our results were similar to those of a previous study in which Salmonella was selected by ciprofloxacin (Wang et al., 2009). This suggests that the expression of invasion-related genes is tightly regulated by environmental conditions such as antimicrobial exposure. Furthermore, we demonstrated that the virulence genes were highly stable in Salmonella during selection with ceftriaxone, in contrast to partial or total loss of the pathogenicity islands in uropathogenic E. coli induced by quinolones (Soto et al., 2006). More in-depth experiments such as growth rate, adherence, and invasion assays have further confirmed the decline of invasion. Compared with a decreased invasiveness of 55.5-fold (Fabrega et al., 2009) after selection by fluoroquinolones in Salmonella Typhimurium mutants, the ceftriaxone-resistant strains declined 34.9-fold. In addition, the Salmonella Typhimurium strains passaged without ceftriaxone had no significant decrease in invasion. Additionally, the percentage of adherence also decreased from 37.8 to 5.06 (7.47-fold) after antibiotic selection. The above data showed that there was a decline of the overall level of Salmonella Typhimurium virulence properties (adherence and invasion) in ceftriaxone-resistant strains.
It is noteworthy that overexpression of arcA, arcB, and tolC after selection with ceftriaxone were 4.0-, 6.8- and 5.4-fold higher, respectively. In addition, ESBLs-producing genes were not detected before and after the selection in the non-ESBLs Salmonella Typhimurium isolates, and the resistance phenotype observed after ceftriaxone selection was thus mainly due to an increased expression of the ArcAB-TolC efflux pump. ArcAB-TolC overexpression is one of the most well-known non-enzymatic resistance mechanisms to quinolones and β-lactam (Li and Nikaido, 2009; Webber et al., 2005). We also sequenced the efflux pump regulatory loci, including acrR, ramR, soxRS, and marRAB. Interestingly, only a silent mutation (T423C) was commonly found in acrR. The encoded product of acrR plays an important role in negative regulation of the ArcAB-TolC efflux pump system. Deletions or point mutations in acrR gene may alter the amino acid sequences of AcrR, resulting in overexpression of ArcAB-TolC (Corvec et al., 2008; Olliver et al., 2004; Webber et al., 2005). However, the detected T423C change in acrR in this study is a silent mutation, which does not alter the encoded amino acid sequences of AcrR. It is unknown if and how this mutation affects the function of acrR and the expression of the efflux pump. It has been reported that a silent mutation occurring in a rare synonymous codon, which requires the use of a rare tRNA for translation, might cause pausing during translation and lead to a change in folding of the nascent protein (Makoff et al., 1989). However, the T423C mutation changes the codon from GTT to GTC, both of which have similar usage frequencies in Salmonella Typhimurium (16.8 and 16.2, respectively) by Codon Usage Database (
It has been reported that the AcrAB-TolC efflux pump of Salmonella Typhimurium plays a role in pathogenesis, and the tolC knockout mutant poorly adheres to both human embryonic intestine cells and mouse monocyte macrophages (Buckley et al., 2006). The deletion of tolC led to down-regulation of Salmonella pathogenicity island-1 (SPI-1) genes related to the type III secretion system-1 (TTSS-1), which was required for Salmonella invasion (Virlogeux-Payant et al., 2008). acrB mutants could reduce the adhesion or invasion, although the effect is still debatable (Baucheron et al., 2005; Buckley et al., 2006). In this study, the up-regulation of the AcrAB-TolC efflux pump and decreased invasion and growth rates were observed in the Salmonella mutants. The increased expression of the efflux system may be required for the elevated antibiotic resistance, but may not be directly linked to reduced virulence. It is possible that during the stepwise antibiotic selection, other unidentified changes occurred in the mutant strains, which modulate the physiological state in Salmonella, rendering the mutant strains less virulent. This speculation awaits further confirmation with additional studies.
Conclusion
The results in this study revealed that stepwise selection of Salmonella Typhimurium with ceftriaxone resulted in the development of cephalosporin-resistant mutants, which showed reduced invasion into epithelial cell and decreased growth rates. The acquisition of antibiotic resistance is correlated with the overexpression of the AcrAB-TolC efflux system, while the reduced invasion is correlated with the decreased expression of the virulence genes invA and stn in the Salmonella Typhimurium mutants. These results suggest that antibiotic selection not only reduces the susceptibility of Salmonella to antibiotics, but also modulates the virulence properties of this pathogenic organism.
Footnotes
Acknowledgments
We are grateful to Professor Qijing Zhang (Iowa State University, Ames) for critically reviewing the manuscript. This work was supported by the National Science Fund for Distinguished Young Scholars (grant 31125026), the Special Fund for Agro-Scientific Research in the Public Interest (grant 201203040), and the National Natural Science Foundation of China (grant U0631006).
Disclosure Statement
No competing financial interests exist.