
Abstract
A robust semiquantitative method for measuring the colonization potential of O157 enterohemorrhagic Escherichia coli (EHEC) strains was developed combining an established ex vivo model infection system, bovine in vitro organ culture, with detection of bacteria attached to tissue sections by immunofluorescent assay (bIVOC-IFA) using Quantum dot® nanocrystal technology. The method was tested on ten O157 strains chosen to reflect a diversity of genotypes found in New Zealand based on the novel polymerase chain reaction–binary typing (P-BIT) system. High- and low-colonizing EHEC O157 strains were identified using bIVOC-IFA, with the highest colonizing strain belonging to the pulsed-field gel electrophoresis type most commonly identified from New Zealand beef meat. Furthermore, all of the toxigenic O157 strains exhibiting a low-colonizing phenotype were closely related, belonging to the same P-BIT genotype cluster. Future use of this method to characterize EHEC strains could provide valuable information for risk assessment and risk management interventions aimed at improving food safety along the beef farm to fork continuum.
Introduction
Ruminants are an important EHEC reservoir, with contamination of associated food products by fecal material containing EHEC being an important vehicle for human infection (Lake et al., 2001). A study in Scotland found that approximately 11.2% of cattle carry EHEC O157 at the time of slaughter (Ogden et al., 2004).
These figures may be even higher in the United States, with one study estimating the prevalence of E. coli O157 on the hides of cull cattle at slaughter to be 46.9% (Brichta-Harhay et al., 2008). Since cattle carry the organism asymptomatically, and carriage can be widespread, control of EHEC O157 on the farm has been difficult and challenging.
An epidemiological model incorporating prevalence data on farms and the heterogeneous fecal shedding rates of E. coli O157 by cattle has determined that 80% of transmission amongst cattle is the result of bacterial shedding by 20% of the most infectious individuals (Matthews et al., 2006). Therefore, control strategies aimed at reducing bacterial loads in these ”super shedder” cattle offer the best potential to reduce the prevalence of EHEC in the beef supply (Chase-Topping et al., 2008; Fairbrother et al., 2006; Stevens et al., 2002). How some animals develop persistent EHEC infection at the terminal rectum and become ”super shedders” is not well understood (Chase-Topping et al., 2005). Potentially, individual EHEC strain characteristics are a factor. One epidemiological study in Scotland has identified a particular O157 phage type (PT), PT 21/28, as a risk factor for super-shedding and severe human morbidity (Chase-Topping et al., 2007; Lynn et al., 2005).
The bIVOC model infection system has previously been used to quantify colonization levels of EHEC O157, with bovine spiral colon and jejunum explants, using a procedure that involved dislodgement of bacteria from the surface of the tissue and enumeration by culture or immunoflourescent detection (Cobbold et al., 2004; Lowe et al., 2009). Although quantitative, this method does not allow for a visual assessment of the quality of the bovine tissue used or the destruction to the mucosal surface that may occur due to the interaction of the bacteria with the tissue. The bIVOC method has been used in combination with immunofluorescent detection of EHEC to characterize the interaction of the bacteria with the tissue surface of bovine terminal rectal explants (Baehler et al., 2000, 2002; Girard et al., 2005), but thus far immunofluorescent detection has not been used to quantify colonization or compare adherence amongst O157 strains. In this study, we developed a semiquantitative method to assess colonization of strains of interest using the bIVOC model infection system in conjunction with immunofluorescent detection of bacteria (bIVOC-IFA) using Quantum dot (Qdot®) nanocrystal technology, a recently described, highly photostable fluorophore (Hotz, 2005). We tested the utility of the culture-independent bIVOC-IFA method with a set of strains chosen to represent a range of O157 genotypes found within New Zealand, as determined by the novel Shiga-toxigenic Escherichia coli (STEC) polymerase chain reaction–binary typing (P-BIT) system (Brandt et al., 2011).
Materials and Methods
Bacterial strain preparation
Bacterial strains were obtained from the New Zealand Culture Collection and the Enteric Reference Laboratory (National Centre for Biosecurity and Infectious Diseases, Wallaceville, New Zealand) (Table 1). EHEC genotype (stx1, stx2, stx2c, stx2d, eaeA, and ehxA) was determined by polymerase chain reaction (PCR) using previously described methods (Paton et al., 1998; Wang et al., 2002). Pulsed-field gel electrophoresis (PFGE) XbaI typing, STEC P-BIT typing, and clustering for the O157 stains used in this study have been previously published (Brandt et al., 2011). Strains were streaked for single colonies from a frozen glycerol stock onto Luria broth (LB) agar plates and grown overnight at 37°C. After incubation, single colonies were inoculated into LB and grown to stationary phase overnight shaking at 37°C. Bacterial cells were pelleted by centrifugation at 4,000 x g for 10 min and resuspended in fresh LB. The final inoculum applied to each tissue biopsy was approximately 108 colony-forming unit (CFU), the minimum dose required to produce colonization by the lowest colonizing strain NZRM3614; this dose is within the range (103 to 108 CFU) used previously to compare O157 strain colonization in a bIVOC adherence assay (Lowe et al., 2009) and is less than the infectious dose used to compare colonization of different O157 strains in cattle using rectal application of bacteria (Sheng et al., 2006).
stx2 allele indicated in subscript parenthesis.
As published in Brandt et al. (2011).
[ ] indicates negative by latex agglutination, but positive by polymerase chain reaction.
Most prevalent PFGE Xba1 type isolated from New Zealand meat.
Non-toxigenic lab strain of O157:H7.
PFGE, pulsed-field gel electrophoresis; STEC, Shiga-toxigenic Escherichia coli; P-BIT, polymerase chain reaction–binary typing.
bIVOC tissue preparation and infection
Explants from the terminal rectum of 2–3-year-old beef cattle (Hereford and Angus breeds) were prepared using previously described methods (Baehler and Moxley, 2000; Girard, 2007). Briefly, fresh biopsies from the last centimeter of the terminal rectum were excised, rinsed in cold, sterile phosphate-buffered saline (PBS), and stored in Roswell Park Memorial Institute (RPMI) 1640 complete medium on ice during transport to the laboratory. RPMI 1640 complete medium contained 10% fetal bovine serum (heat inactivated), 0.25% lactalbumin hydrolysate, 0.2 μg/mL hydrocortisone, 0.1 μg/mL insulin, 75 mM β-mercaptoethanol, 2 mM
Immunostaining
Tissue sections on glass slides were deparaffinised by incubation at 60°C until the wax had melted. All other procedures were conducted at room temperature unless otherwise stated. Slides were washed twice in SafSolvent (Ajax Finechemicals, Auckland, New Zealand) for 5 min. Slides were rehydrated in decreasing ethanol concentrations (100%, then 95%, then 70%) for 3 min in each solution and then washed twice in PBS for 5 min. Slides were blocked in 6% bovine serum albumin/0.1% Triton X-100 in PBS for 1 h. Primary anti-serum, Difco rabbit anti-O157 serum (Fort Richard Laboratories, Auckland, New Zealand), was diluted 1:100 in blocking solution/10% normal rabbit serum and applied to slides for 1 h. Slides were then washed three times in PBS. The secondary antibody, goat anti-rabbit IgG conjugated to Qdot® 655 (Invitrogen Corp., Carlsbad, CA), was diluted 1:500 in blocking solution/10% normal goat serum/5 μg mL−1 Hoechst 33342 (Invitrogen Corp.) and applied to slides for 1 h. Slides were washed three times in PBS and mounted with Vectashield (Vector Labs, Burlingame, CA) under a no. 1 glass coverslip sealed to the slide with nail polish. Qdot® fluorophore-conjugated secondary antibodies were used to provide additional brightness and stability during confocal imaging. We did not observe any photobleaching during image acquisition and found that tissue staining was stable for at least 6 months when slides were stored in the dark at 4°C.
Semiquantifying bacterial colonization using confocal microscopy
Tissue sections were analyzed using an inverted Leica TCS SP5 confocal laser scanning microscope (CLSM) with LAS AF imaging software (Leica Microsystems, Wetzlar, Germany). Bacterial cells were visualized using a 100× NA 1.2 oil immersion lens with 2× zoom using excitation 488 nm and emission of 630–690 nm. The majority of bacterial cells attached to the surface of the terminal rectal tissue were enumerated by eye along the entire length of the tissue section. In the case of the high-colonizing strains in particular, bacterial clumps were observed, which were more difficult to count by eye. These areas were photographed and as many bacteria as possible were counted by digital zooming of electronic images. As this method is semiquantitative, failure to resolve all of the bacteria in the images did not change the placement of a strain as a high or low colonizer. Enumeration was performed from three explants per strain taken from three separate animals.
The total number of bacterial cells was then normalized to the surface length (mm) of the tissue section. The surface length of a given tissue section was determined by using a standard metric ruler and light box. Single bacteria were counted individually through the microscope ocular. Bacteria growing in clumps were counted by generating a series of z-stack images of width 0.5 μm. The tissue integrity of each immunostained sample was assessed by differential interference contrast (DIC) microscopy; only sections with comparable tissue integrity to uninfected control samples were used for bacterial cell enumeration and subsequent comparison between bacterial strains. A representative image of an immunostained section from a high-colonizing EHEC O157 strain is presented in Figure 1.
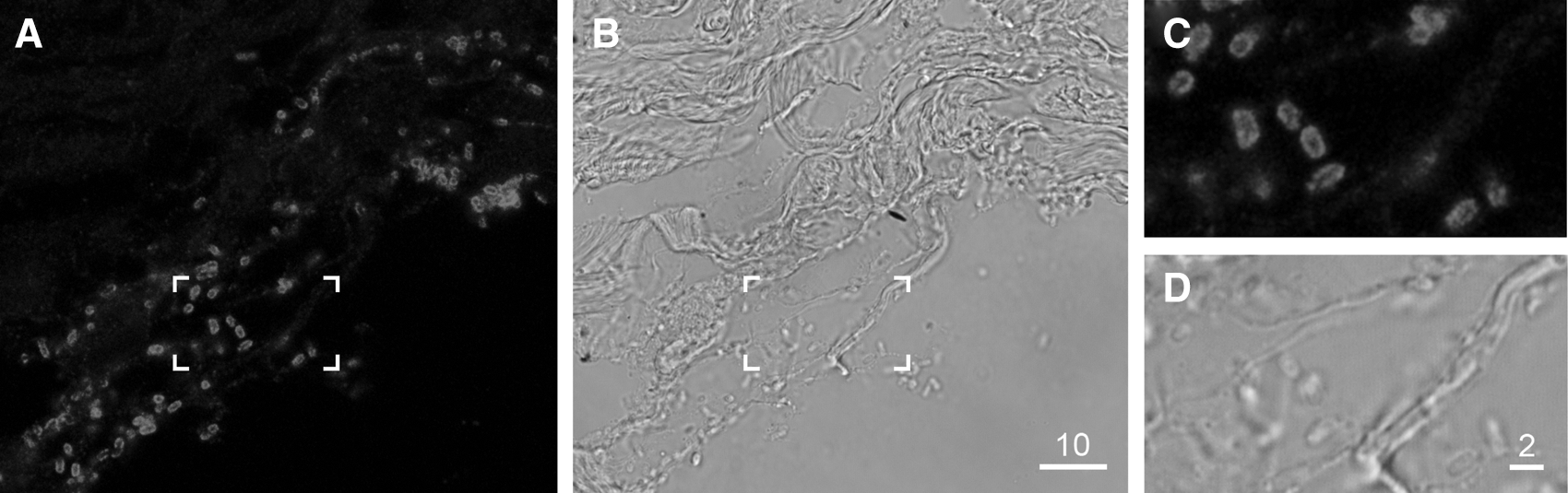
Representative image of O157 enterohemorrhagic Escherichia coli (strain ERL04-3476) colonizing the surface of a bovine terminal rectum explant. Bacteria were detected by immunostaining using a primary antisera recognizing the O157 O-antigen and a secondary antibody conjugated to Quantom dot (Qdot®) 655. Confocal optical sections were collected at 0.5-μm intervals with a 100× NA 1.2 oil immersion lens using 488-nm excitation with fluorescence collected at 630–690 nm. A single representative section is shown.
Statistical analysis
Strains were separated into two groups: high colonizers (>50 bacteria per mm tissue section in two or more experiments) and low colonizers (<50 bacteria per mm tissue section in all experiments). The Mann-Whitney one-tailed U-test was used to determine if the attachment and growth of low-colonizing strains was less than high-colonizing strains. This non-parametric test was used due to the difference in variances of the two groups, due to inter-experimental variation, and to avoid normality assumptions.
Results
The colonization potential of 10 EHEC O157 strains was quantified using bIVOC-IFA (Fig. 2). Bacterial strains were selected based on diverse origin of isolation and genotype, using the novel P-BIT typing system in an effort to test a range of strains found within New Zealand (Table 1) (Brandt et al., 2011). The strain set included the most commonly isolated PFGE type found in New Zealand beef products (Xb0040; personal communication, Dr. Brent Gilpin) as a potential high-colonizing strain, and a highly passaged attenuated (non-toxigenic) O157 lab strain (NZRM3614) as a potential low-colonizing strain.
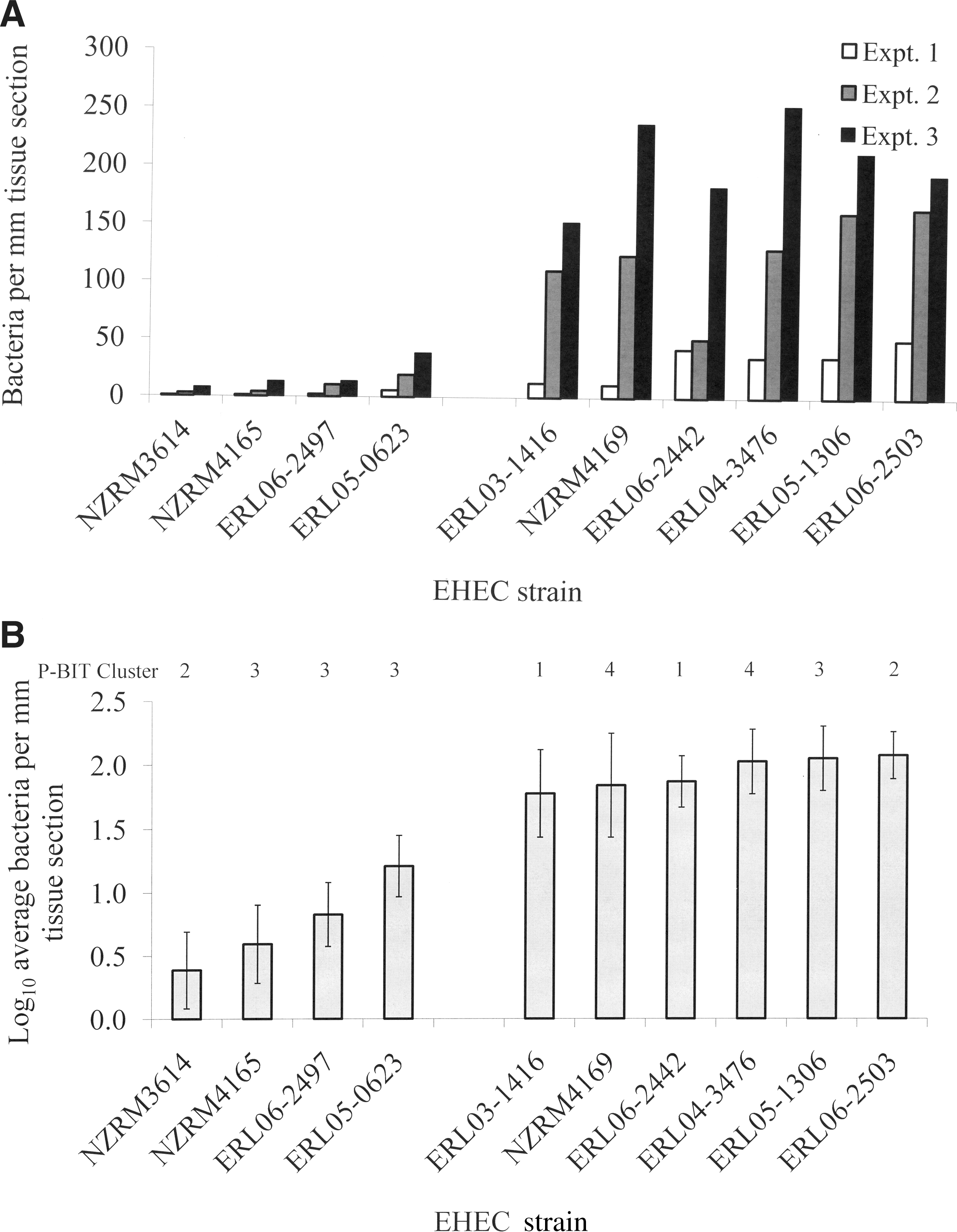
Semiquantification of colonization levels of bovine terminal rectum explants by different O157 strains using bovine in vitro organ culture, with detection of bacteria attached to tissue sections by immunofluorescent assay (bIVOC-IFA).
The bIVOC-IFA model infection system was robust enough to separate the O157 strain set into two groups (p<0.001 by Mann-Whitney one-tailed test): a group of low-colonizing strains (n=4) and a group of high-colonizing strains (n=6; Fig. 2A). There was a one log10 difference in bacterial numbers attached to the mucosal surface of a representative tissue section between the low-colonizing group (strain average 0.75±0.15 log10 CFU per mm tissue section surface) and the high-colonizing group (strain average 1.9±0.10 log10 CFU per mm tissue section surface; Fig. 2B). The strain with the lowest colonization levels was an attenuated non-toxigenic lab strain (NZRM3614) and the highest colonization levels were measured for the O157 strain (ERL06-2503) with the PFGE type most commonly isolated from New Zealand beef (Xb0040). Along with the non-toxigenic lab strain, the low-colonizing group of strains consisted of three O157 strains from the same O157 P-BIT cluster, cluster 3, of strains with the distinguishing feature of possessing both the stx1 and stx2 genes.
Discussion
A semiquantitative method called bIVOC-IFA was developed to assess the colonization potential of O157 strains by combining an established ex vivo model infection system with immunofluorescent detection of bacteria attached to tissue sections. The follicle-rich region of the bovine terminal rectum was used because this region of the gastrointestinal tract is a location where EHEC O157 in cattle naturally persists and because terminal rectal explants have previously been used to study the interactions of this serotype with host gut tissue (Girard et al., 2007; Lim et al., 2007; Naylor et al., 2003, 2005). The low-colonizing strains identified by the bIVOC-IFA colonization assay included the attenuated non-toxigenic lab strain and strains carrying both stx1 and stx2 genes (P-BIT cluster 3). All O157 carrying stx2 alone displayed high colonization in the bIVOC-IFA assay. These results are consistent with studies of the prevalence of stx genes in bovine O157 isolates, which have reported the presence of stx1 much less frequently (<20%) than the presence of stx2 (100%) (Alam et al., 2006; Lefebvre et al., 2009; Paiba et al., 2002). O157 strains carrying both stx1 and stx2 have also been reported to be less virulent in humans and are less cytotoxic in in vitro cell assays and in in vivo neurotropic assays in piglets than strains carrying stx2 alone (Diarra et al., 2009; Donohue-Rolfe et al., 2000; Ethelberg et al., 2004; Griffin et al., 1991; Lefebvre et al., 2009; Rivas et al., 2006).
The one inconsistency in our results is the stx1- and stx2-carrying strain ERL05-1306 (P-BIT cluster 3) which exhibited a high colonization phenotype in our assay. This is an intriguing result because this strain was chosen for the study because it is the only non-P-BIT cluster 4 strain that we have identified thus far which carries the stx2c allele (Brandt et al., 2011). Similar to ERL05-1306, both stx2c-carrying (PBIT cluster 4) O157 strains that were tested in our bIVOC-IFA model infection system displayed a high colonization phenotype. This is consistent with a number of studies that have identified stx2c as the dominant subtype associated with bovine isolates (Oseki et al., 2002; Ding et al., 2011; Franz et al., 2012), which would suggest a propensity for these strains to colonize cattle. Furthermore, the PFGE type most commonly isolated from New Zealand beef products, represented by strain ERL06-2503, displayed the highest colonization levels in the bIVOC-IFA colonization assay. Taken together, these preliminary results suggest that our model bovine colonization system can identify “low-risk” strains from “high-risk” strains, in terms of prevalence amongst cattle.
Despite inter-experimental variation (Fig. 2A), which may have been attributed to a number of parameters (e.g., quality of the bovine tissue sample, use of outbred animals, quality of tissue sectioning, and/or the strength of immunostaining), partitioning of strains into low- and high-colonizing groups was achieved using the bIVOC-IFA method. Use of a representative section from the middle of the explant was sufficient to discriminate between the two phenotypic groups, as the bIVOC-IFA method was capable of identifying the attenuated lab strain and the most prevalent O157 phage type found in New Zealand beef products as the lowest and highest colonizing strains, respectively.
The benefit of using bIVOC-IFA, as opposed to previously published methods used to enumerate E. coli colonization on bovine tissues (Cobbold et al., 2004; Lowe et al., 2009), is that bIVOC-IFA does not require dislodging bacteria from the surface of the tissue, which is advantageous as the samples could also be used to assess differences in colonization patterns and tissue pathology. More importantly, bIVOC-IFA allows visual comparison of tissue integrity between samples, which is important when making comparisons between bacterial strains as a low-colonizing phenotype could be misinterpreted due to poor quality of the tissue explant or severe destruction of the tissue surface mucosa during infection. The bIVOC-IFA method can be completed within 2 working days, which is similar to culture enumeration methods that require overnight incubation. Furthermore, the bIVOC-IFA method could be modified for high-throughput analysis by using a qualitative assessment of slides viewed at a lower (e.g., 10×) magnification with a standard fluorescent microscope.
Conclusion
This work describes a semiquantitative method for enumerating the colonization potential of a major foodborne pathogen, using an established ex vivo model infection system in a host animal responsible for zoonotic infections. The bIVOC-IFA method could be used by the beef industry to target potential interventions to farms harboring EHEC O157 super-shedding strains, those identified as high-colonizing strains in the bIVOC-IFA assay. The results presented in this study are indicative of the potential for bIVOC-IFA to be used in this manner; however, further validation of the method would first be required using a well-defined set of EHEC O157 strains with known prevalences and shedding characteristics in New Zealand cattle. In addition to application within the beef industry, this method could be used as a model system to compare environmental and clinical EHEC O157 strains in regards to their potential to colonize a bovine host, providing valuable strain-based information for risk assessment and modeling aimed at improving food safety.
Footnotes
Acknowledgments
We would like to thank South Pacific Meats Ltd. (Malvern, Christchurch, New Zealand) for providing bovine colonic tissue and Carolyn Nicol (ESR, New Zealand) for use of select EHEC strains. We would like to thank Drs. Manfred Ingerfeld and David Collings (University of Canterbury, Christchurch, New Zealand) for technical assistance in generating the CLSM images, Jennifer Lucas (Gribbles Veterinary, Christchurch, New Zealand) for tissue sectioning, and Beverley Horn (ESR, New Zealand) for help with statistical analysis of the data. This work was enabled by capability funding provided by the New Zealand Ministry of Research Science and Technology.
Disclosure Statement
No competing financial interests exist.