
Abstract
Shigatoxigenic Escherichia coli (STEC) cause serious foodborne infections that lead to diarrheal disease and sequelae worldwide. In Burkina Faso, West Africa, STEC strains from environmental and human sources have not been isolated and characterized before. In this study, 21 STEC strains were isolated from food samples of animal origin and human feces using colony hybridization of the Shiga toxin gene stx. The STEC strains belonged to 15 different serotypes, including O43:H2, O8:H-, and O2:H2. All strains were positive for stx1 and 10 also for stx2 . The most common stx1 subtype was stx1a , and the most common stx2 subtype was stx2b . In five strains, stx2 subtypes stx2a and/or stx2c , which were previously associated with hemolytic uremic syndrome, were present. Some of the strains possessed the gene saa, encoding autoagglutinating adhesin. None of the strains possessed the gene eae, encoding intimin. Two STEC strains carried also an enterotoxigenic E. coli–associated gene estIa, encoding heat-stable enterotoxin. The STEC isolated from food in Burkina Faso are potentially pathogenic for humans based on the virulence gene combinations that they possess and phenotypes that they express.
Introduction
S
Shiga toxin (Stx) is the main virulence trait of STEC. There are two types of Shiga toxin, Stx1 and Stx2, encoded by the genes stx1 and stx2 , respectively, and both of them have several subtypes. The following designations for the subtypes have been suggested: for stx1 subtypes stx1a , stx1c , and stx1d ; and for stx2 subtypes stx2a , stx2b , stx2c , stx2d , stx2e , stx2f , and stx2g (Scheutz, 2009). STEC strains can express either one or several Stx1 and/or Stx2 subtypes (Nataro and Kaper, 1998; Brett et al., 2003). Of the main toxin genes, stx2 is more frequently associated with hemolytic uremic syndrome (HUS) than stx1 alone or stx1 and stx2 together (Friedrich et al., 2002). In particular, subtypes stx2a and stx2c have been associated with HUS (Eklund et al., 2002; Persson et al., 2007).
Some STEC strains have the ability to cause attaching and effacing (A/E) lesions on the surface of the intestinal epithelial cells (Nataro and Kaper, 1998). This feature is mediated by a protein called intimin, which is encoded by the gene eae located in the pathogenicity island Locus of Enterocyte Effacement (LEE). These strains are thought to cause more severe infections in humans than intimin-negative STEC (Paton and Paton, 1998). However, intimin-negative STEC have been isolated from HUS patients (Brett et al., 2003; Galli et al., 2010). Prevalence of eae is significantly more common among STEC isolated from humans than among STEC from cattle feces and beef (Pradel et al., 2000; Mora et al., 2005). Certain STEC serogroups carry eae more commonly than others. For example, in one study, all the isolated STEC O157:H7 strains of human, cattle, and beef origin carried eae, whereas few of the non-O157 strains did (Mora et al., 2005). STEC often possesses other putative virulence factors, too. The gene hlyA (often designated as ehxA) required for the production of enterohemolysin is common in STEC isolated from beef and cattle feces, but also in STEC from hospitalized children (Pradel et al., 2000). STEC autoagglutinating adhesin (Saa), encoded by the gene saa, is found from LEE-negative strains of human, cattle (Galli et al., 2010), and food (Herold et al., 2009) origin. Saa in intimin-negative STEC possibly helps the bacteria to attach on the intestinal epithelial cells (Paton et al., 2001), and it has been identified from intimin-negative STEC isolated from HUS patients (Herold et al., 2009; Galli et al., 2010).
In this study, we isolated and characterized STEC strains isolated from raw meats and intestines sold at the common markets and in human stool samples from Burkina Faso, West Africa. We have previously reported that up to 43% of the beef products (34% of the beef and 43% of the bovine intestines) and 20% of the mutton sold at the open markets in Ouagadougou, the capital of Burkina Faso, contained STEC or other DEC (Kagambèga et al., 2012). However, STEC was found to be rather rare in the Burkinabe diarrheal children (2%), although the overall DEC infection rate was 45% (Bonkoungou et al., 2012).
The aim of this study was to evaluate the virulence potential of the isolated STEC. This article describes the pheno- and genotypic characteristics of STEC strains found in this geographical area and, thus, provides useful background information for future food safety and public health surveillance programs.
Materials and Methods
Samples and isolation of STEC strains
Retail meats and intestines (120 samples) sold at open markets and human stool samples from children with (471 samples) or without diarrhea (187 samples) were collected in Burkina Faso during 2008–2010 as described by Bonkoungou et al. (2012) and Kagambèga et al. (2012). Briefly, the beef, bovine intestine, mutton, and chicken samples were collected from four markets in Ouagadougou (Bendogo, Koulouba, Zogona, and Dassasgho). Stool samples were from hospitals in the city of Ouagadougou and in rural areas of Boromo and Gourcy. For meat and intestine samples, 25 g of sample was added into 225 mL of Buffered Peptone Water (Liofilchem, Teramo, Italy), homogenized, and streaked onto Müller Hinton agar plates (Liofilchem) to grow. Human stool samples were cultured directly onto Sorbitol-MacConkey agar plates (SMAC; Lab M, Bury, Lancashire, UK). The bacterial mass was collected from the plates for storage. For polymerase chain reaction (PCR) analysis, the bacterial mass was re-cultured onto Cystine Lactose Electrolyte Deficient agar plates (meat and intestine samples; Lab M) or SMAC agar plates (stool samples). Multiplex PCR was used to detect DEC virulence genes in the mixed bacterial cultures as previously described (Bonkoungou et al., 2012; Kagambèga et al., 2012). Multiplex PCR detected the following 16 genes of different DEC pathogroups (from the longest gene fragment to the shortest): uidA, pic, bfpB, invE, hlyA, elt, ent, escV, eae, ipaH, aggR, stx1 , stx2 , estIb, estIa, and astA (Antikainen et al., 2009). Forty-one mixed bacterial cultures were found to be stx-positive, 33 from meat or intestine samples and eight from stool samples, and they were subjected to colony hybridization to isolate STEC cultures. Before hybridization, the cultures were grown on SMAC agar at +36°C for 18 h. The colonies were replicated onto the Nylon Membranes for Colony and Plaque Hybridization (Roche Diagnostics GmbH, Mannheim, Germany). Lysis of the bacterial cells, and DNA denaturation and binding to the nylon membrane were performed according to the manufacturer's instructions. For hybridization, 4 ng of a digoxigen-labeled DNA probe specific for stx1 or stx2 (Statens Serum Institut [SSI], Copenhagen, Denmark) was used in combination with DIG Nucleic Acid Detection Kit (Roche Diagnostics, Indianapolis, IN). The colonies that appeared to be stx-positive by hybridization were re-cultured on SMAC and verified as STEC using the multiplex PCR.
Strain characterization
In addition to the 16 genes included in the multiplex PCR, the presence of saa was defined using a multiplex PCR (Paton and Paton, 2002) or a gene probe at SSI Diagnostica typing service (
Results
Altogether, 21 STEC strains were isolated from the 41 stx-positive mixed bacterial cultures from beef, mutton, and bovine intestines (20 strains) and the human feces samples (one strain from a child with diarrhea). All strains carried stx1 and 10 also stx2 (Table 1). The strains with both stx genes were, in most cases, of bovine origin. The stx subtypes stx1a , stx1c , stx1d , stx2a , stx2b , stx2c , and stx2d were present in STEC of meat origin and stx1c in STEC from the human feces sample in Burkina Faso. The most common stx1 subtypes were stx1a (nine strains), followed by stx1c (eight strains) and stx1d (four strains). The most common stx2 subtypes were stx2b (four strains), followed by stx2a (three strains), stx2c (three strains), and stx2d (one strain). One of these strains possessed both stx2a and stx2c . Among the strains possessing both stx1 and stx2 genes, the most common subtypes were stx1c and stx2b (four strains). None of the strains possessed subtypes stx2e , stx2f , or stx2g . All the strains were eae-negative. The gene hlyA was present in 15 strains, and seven of these were also positive for saa. The most common virulence gene profiles were stx1c together with hlyA (four strains) and stx1d alone (four strains; Table 2). The genotype stx1c together with hlyA was found from both the human and three meat isolates.
Production of β-glucuronidase.
Hemolysis after 24-h incubation on washed sheep blood agar with Ca2+.
Hemolysis after 4-h incubation on washed sheep blood agar with Ca2+.
Susceptible to the tested 12 antimicrobials: ampicillin, chloramphenicol, streptomycin, sulphonamide, tetracycline, trimethoprim, ciprofloxacin, gentamicin, nalidixic acid, cefotaxime, mecillinam, and imipenem.
Intermediate susceptibility to streptomycin only and susceptible to the remaining 11 antimicrobials.
One strain was from the human feces, and three strains were from meats.
The STEC strains belonged to 15 different serotypes, with O43:H2 being the most common (Table 1). The bovine, mutton, and human strains did not share the same serotypes. Two STEC strains of the serotype O2:H2 from bovine sources possessed an ETEC-associated gene estIa, encoding heat-stable enterotoxin Ia. All the hlyA-positive strains were enterohemolytic on washed sheep blood agar. All the strains were susceptible to the tested antimicrobials, except that one strain from mutton and one from bovine intestine had lowered susceptibility to streptomycin. Sorbitol was fermented and PGUA produced by 18 strains. The human isolate produced Shiga toxin in the dilution 1:8.
PFGE revealed that strains of the same serotype and carrying the same virulence genes were genetically similar (Fig. 1 and Table 1). An exception was made by the two strains of serotype O2:H2, which had the same virulence gene profile but different PFGE profiles.
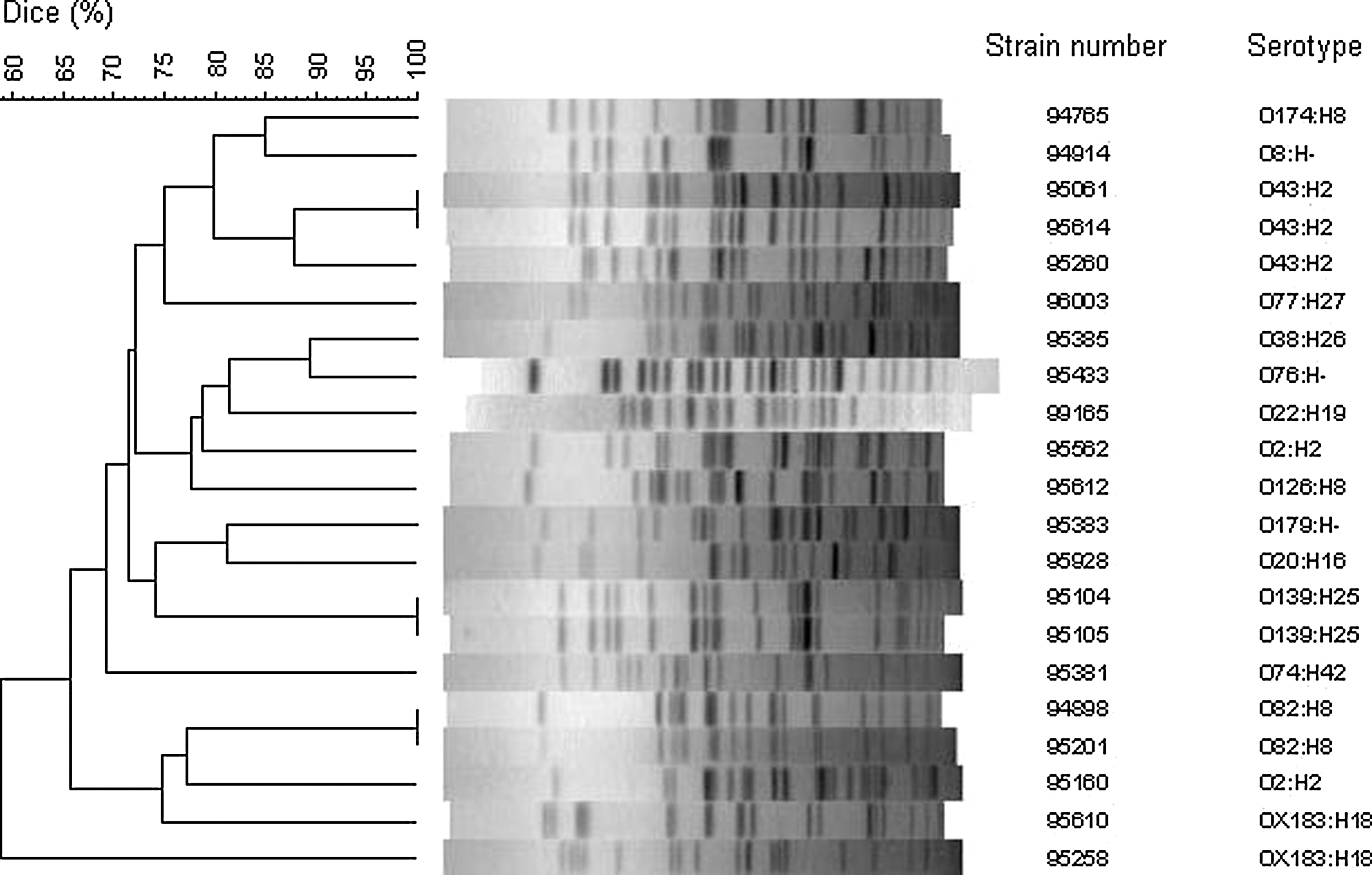
Unweighted Pair Group Method with Arithmetic Mean (UPGMA) dendogram of pulsed-field gel electrophoresis (PFGE) profiles obtained using XbaI digestion. Similarities were calculated using Dice algorithm.
Discussion
We characterized the virulence properties and estimated the virulence potential of 21 STEC strains from Burkina Faso by phenotyping and genotyping. The Burkinabe food-derived STEC carried stx1 alone or in combination with stx2 . Our previous PCR detection results showed that stx1 was more common (23%) than stx2 (18%) in raw meat products, but 6% of the stx-positive samples were positive for stx2 only (Kagambèga et al., 2012). Also in Australia, stx1 alone or in combination with stx2 was found to be more prevalent than stx2 alone in STEC from human and ruminant feces and meat (Brett et al., 2003). However, stx2 alone was found to be more common than stx1 alone in non-O157 STEC from cattle feces and meat products in Brazil (Irino et al., 2005), in Spain (Mora et al., 2005), and in Germany (Martin and Beutin, 2011). This probably indicates epidemiological variability also in the occurrence of different serotypes not only different stx virulence genes between different regions.
The most common stx1 subtypes were stx1a and stx1c , whose prevalence was almost equal. STEC carrying stx1a has previously been found from different foods such as beef, cow milk, and rabbit meat (Martin and Beutin, 2011). The gene stx1c has previously been associated with sheep and mutton (Brett et al., 2003; Martin and Beutin, 2011). Likewise, we found STEC carrying stx1c in all our three mutton isolates, but also in four bovine isolates. STEC of the serotype O174:H8 with stx1c has previously been isolated from a HUS patient, sheep, and cattle feces (Koch et al., 2001; Brett et al., 2003). The human isolate carried stx1c , which was also carried by the meat isolates. As has also been reported by others (Martin and Beutin, 2011), stx1d was the least prevalent of the stx1 subtypes present in the meat isolates in our study.
Among the Burkinabe STEC, the most common stx2 subtype was stx2b , which was found in the strains of both bovine and ovine origin. The gene stx2b has previously been found to be common in sheep and goat meat and milk (Martin and Beutin, 2011). Two Burkinabe strains possessed stx2a and two stx2c , and one strain possessed both of them. The genes stx2a and stx2c have been identified as the most prevalent stx2 subtypes in STEC isolated from ground beef in the United States (Bosilevac and Koohmaraie, 2011). In addition, subtypes stx2a and stx2c have been associated with HUS previously (Eklund et al., 2002; Persson et al., 2007). Both stx2a and stx2c were detected only from bovine sources in Burkina Faso. The gene stx2d was detected in one beef strain. STEC harboring stx2d has previously been isolated from diarrheal calves (Nguyen et al., 2011), from patients with bloody (Persson et al., 2007) and non-bloody diarrhea, and from asymptomatic carriers (Friedrich et al., 2002).
All the strains were eae-negative. Intimin, encoded by eae, has been thought to be necessary for STEC to be able to cause disease in humans (Kaper et al., 2004). However, intimin-negative strains have caused severe disease (EFSA, 2011), and there are other potential virulence factors possibly replacing intimin in the intimate adhesion needed for the full virulence. The saa gene together with the ability to produce enterohemolysin, as shown by the in vitro test, could make the seven found saa-carrying food isolates a potential cause of HUS (Law and Kelly, 1995; Aldick et al., 2007).
Two STEC strains of the serotype O2:H2 of the bovine origin possessed an ETEC-associated gene estIa encoding heat-stable enterotoxin Ia. DEC are able to acquire virulence genes via horizontal gene transfer from other pathogroups leading to intermediate pathogroups (Müller et al., 2007) and possibly to the emergence of more pathogenic strains (EFSA, 2011). STEC- and ETEC-associated virulence genes have also previously been identified to exist together in the same strain (Müller et al., 2007; Fratamico et al., 2008; Lienemann et al., 2011). Recently, a strain that had STEC- and EAEC-associated virulence properties caused a large outbreak in Germany (EFSA, 2011). In the present study, beef and bovine intestine samples contained the intermediate strains, and bovine gut might be a potential place for gene transfer to occur between E. coli strains.
None of the Burkinabe strains were of serotype O157:H7, nor were they of the other STEC serogroups commonly reported to cause infections in humans: O26, O55, O91, O103, O111, O113, and O145 (Perelle et al., 2004). However, the same serotypes as isolated by us, O8:H-, O38:H26, O74:H42, O82:H8, O126:H8, O174:H8, O179:H-, and OX183:H18 (WHO, 1998; Pradel et al., 2000; Friedrich et al., 2002; Scheutz et al., 2004; Prager et al., 2005; Bielaszewska et al., 2006; Buvens et al., 2010), have previously been isolated from patients suffering from diarrhea, and they also have possessed virulence gene profiles similar to those of our STEC, except our STEC O179:H- was eae-negative, whereas Scheutz et al. (2004) reported eae-positive STEC O179:H-. In addition, STEC of the serotype O2:H2 (WHO, 1998) and O174:H8 (Brett et al., 2003) have previously been isolated from patients with HUS. STEC of the serotype O43:H2 has been isolated from a human sample in Finland, but no clinical data were available (Eklund et al., 2002). To our knowledge, STEC of serotype O22:H19, which was isolated from a diarrheal Burkinabe child, has not been isolated from humans before, only from ground beef (Bosilevac and Koohmaraie, 2011). We did not find any previous reports on STEC of serotype O77:H27 or O139:H25. Serotypes O20:H16 and O76:H- have been isolated from cows (Hussein and Bollinger, 2005) and sheep (Rey et al., 2003), respectively, but not from humans. The serotypes O8:H-, O38:H26, O43:H2, O74:H42, O82:H8, O126:H8, O174:H8, and O179:H-, which have been isolated from diseased humans, have previously been isolated also from cattle and sheep, and from food derived from these animals (Read et al., 1992; Hussein and Bollinger, 2005; Gomes Oliveira et al., 2008; Fernándes et al., 2010; Martin and Beutin, 2011). Thus, the same serotypes of STEC have been found from animal-derived meat from Burkina Faso and from industrialized countries such as Germany (Martin and Beutin, 2011).
Raw meat sold at the open markets in Ouagadougou was found to be heavily contaminated with STEC (Kagambèga et al. 2012). In that study, some of the raw meat samples examined by direct PCR detection were found to contain both Shiga toxin and intimin genes. Now, when studying the isolated STEC, we never found the stx and eae genes present together in the same strain, indicating that the previously studied samples probably contained both STEC and EPEC.
DEC infections in children younger than 5 years of age are common in Burkina Faso (Bonkoungou et al., 2012). However, STEC was not identified as a common pathogen in children suffering from diarrhea (2%). The youngest children do not consume meat, but food for children is prepared in the same environment where raw meat products are handled. Thus contaminated meat can be a source of STEC infections even to the young children. Due to modest medical resources, serious STEC infections and sequelae such as kidney injuries go unreported in Burkina Faso.
Conclusion
Our data show that retail meats in Burkina Faso are a reservoir of STEC strains belonging to diverse serotypes and with various virulence profiles. Based on the pheno- and genotypic characteristics of the isolated STEC strains, they could potentially be a serious health risk to people in Burkina Faso.
Footnotes
Acknowledgments
We thank the personnel of the Bacteriology Unit at the Finnish National Institute for Health and Welfare (THL). The study was funded by the Academy of Finland grant 122600 for the collaboration between THL and CRSBAN/University of Ouagadougou, Burkina Faso.
Disclosure Statement
No competing financial interests exist.