
Abstract
The purpose of this study was to determine the prevalence and characteristics of CTX-M β-lactamases in Escherichia coli among healthy swine and cattle in Korea. A total of 1212 fecal samples obtained from healthy pigs (n=558) and cattle (n=654) were screened for CTX-M–type extended spectrum β-lactamase (ESBL)–producing E. coli isolates. One hundred and twenty-one E. coli that produced ESBL were subjected to phenotypic and genotypic characterization. A high number (120/558, 21.5%) of swine fecal samples showed the presence of CTX-M β-lactamase–producing E. coli compared to cattle samples (1/654, 0.2%). The most predominant CTX-M–type identified was CTX-M-14 (n=82), followed by CTX-M-15 (n=16). Isolates producing CTX-M-3, CTX-M-27, CTX-M-55, and CTX-M-65 were also identified. Overall, the bla TEM-1 gene was associated with CTX-M β-lactamase in 55 E. coli isolates. Transfer of bla CTX-M gene was demonstrated from 76 out of 121 bla CTX-M-positive E. coli isolates to the recipient E. coli J53 by conjugation. Plasmid DNA isolation from the transconjugants revealed a large (90–120 Kb) conjugative plasmid. ISEcp1 and IS903 were detected upstream and downstream of bla CTX-M genes in 117 and 91 E. coli isolates, respectively. Our results demonstrated that a combination of clonal expansion and horizontal transmission is spreading bla CTX-M genes among swine E. coli. The horizontal dissemination of bla CTX-M genes among E. coli was mostly mediated by IncF or IncI1-Iγ plasmids. To the best of our knowledge, this study represents the first report of CTX-M-3, CTX-M-27, CTX-M-55, and CTX-M-65 β-lactamases in bacterial isolates from food animals in Korea. This study revealed that the CTX-M β-lactamase–producing E. coli are widely disseminated among healthy pigs but very rare in cattle in Korea. Increasing prevalence of bla CTX-M genes in intestinal E. coli of food animals is a matter of concern and should be carefully monitored.
Introduction
Humans, via the food chain, ingest a lot of bacteria originating from food-producing animals, which have been recognized as major reservoirs of E. coli–producing CTX-M β-lactamase (Ho et al., 2011). Recently, a high prevalence of CTX-M β-lactamase–producing Enterobacteriaceae has been reported in stool specimens from healthy asymptomatic volunteers in a rural community in Thailand (Sasaki et al., 2010). Thus, there are concerns about the potential spread of the bla CTX-M genes from food animal products to humans through the food chain.
CTX-M β-lactamase has been increasingly reported in E. coli from various food-producing animals worldwide raising a potential threat to public health (Li et al., 2007). The earliest account of CTX-M β-lactamase of food animal origin came from Spain, where a CTX-M-14–producing E. coli was isolated from healthy chickens (Brinas et al., 2003). Since then, E. coli–producing CTX-M β-lactamase has been reported from healthy cattle from Japan (Shiraki et al., 2004) and Hong Kong (Duan et al., 2006), and from sick or healthy cattle from France (Meunier et al., 2006; Madec et al., 2008) and the United States (Wittum et al., 2010). Similarly, CTX-M β-lactamase–producing E. coli strains in pigs have been previously reported from Hong Kong (Ho et al., 2011), China (Liu et al., 2007), Spain (Blanc et al., 2006), and France (Meunier et al., 2006). Moreover, we reported CTX-M-14– and CTX-M-15–producing E. coli from sick cattle and sick pig, respectively, for the first time in Korea (Lim et al., 2009). However, there is no data on CTX-M β-lactamases in E. coli isolates from healthy food animals from Korea, contrary to those in the human field (Lee et al., 2009; Song et al., 2009). This study was therefore, undertaken to determine the prevalence of CTX-M β-lactamase–producing E. coli among healthy swine and cattle, and to characterize the CTX-M β-lactamase–producing E. coli using molecular methods.
Materials and Methods
A total of 1212 unique fecal samples obtained from 558 healthy pigs from 56 farms and 654 healthy cattle from 66 farms located throughout Korea during 2008–2009 were investigated. All farms studied were conventional farms. Swine feces were collected from carcasses of animals from eight different slaughterhouses, whereas cattle feces were collected directly from the farms. The samples were inoculated on MacConkey agar plates supplemented with 2 μg/mL cefotaxime (CTX), and the suspected pink isolates were subsequently subcultured on Chromogenic ESBL agar (BioMerieux, Mstvy-l'Etoile, France). Only one isolate per fecal sample was included in the study. Identification of E. coli isolates were done by standard biochemical tests and confirmed by the Vitek system (BioMerieux, Hazelwood, MO).
ESBL production was detected by double-disc synergy test and was confirmed by the Epsilometer test (Etest) using cefotaxime-cefotaxime-clavulanic acid and ceftazidime-ceftazidime-clavulanic acid strips (AB Biodisk, Solna, Sweden) according to Clinical Laboratory Standards Institute (CLSI) guidelines (CLSI, 2009).
Antimicrobial susceptibility was determined by the standard disc diffusion method on Mueller Hinton agar (BD; Becton-Dickinson, Sparks, MD) in accordance with CLSI guidelines (CLSI, 2009) using commercial discs (BBL; Becton-Dickinson, Cockeysville, MD). The antimicrobial drugs tested were ampicillin, amoxicillin-clavulanic acid, cephalothin, cefotaxime, ceftazidime, cefoxitin, imipenem, chloramphenicol, gentamicin, neomycin, streptomycin, tetracycline, ciprofloxacin, nalidixic acid, and trimethoprim-sulfamethoxazole. Minimum inhibitory concentrations (MICs) of selected antimicrobials were determined by Etest method (AB Biodisk) according to CLSI recommendations (CLSI, 2009). E. coli American Type Culture Collection (ATCC) 25922 was used as quality control strain.
The presence of bla
CTX-M gene was screened as described previously (Batchelor et al., 2005). For CTX-M–positive isolates, group-specific primers for CTX-M-1 and CTX-M-9 families were used to amplify bla
CTX-M genes (Branger et al., 2005). A combination of CTX-M-1G FL-F (Yu et al., 2007) or CTX-M-9G FL-R (Xiong et al., 2007) primers and ISECP1U1 primer (Saladin et al., 2002) were used to amplify and sequence the complete bla
CTX-M genes. Polymerase chain reaction (PCR) amplification and sequencing of entire bla
TEM and bla
SHV genes was done as described previously (Rayamajhi et al., 2008). Sequence analyses and comparison with known sequences were performed with the BLAST programs at the National Center for Biotechnology Information website (
Conjugation was performed by broth mating at 37°C on all CTX-M–positive E. coli isolates using plasmid-free sodium azide–resistant E. coli J53 as recipient as described previously (Tamang et al., 2007). Transconjugants were selected on MacConkey agar plates supplemented with sodium azide (150 mg/L) (Sigma) and cefotaxime (2mg/L). All transconjugants were examined for the presence of β-lactamase genes, insertion sequences (IS) and tested for susceptibility pattern as described above.
Plasmid DNAs were isolated using QuickGene® plasmid isolation system (FUJIFILM Corporation, Tokyo, Japan) according to the manufacturer's instructions. Plasmid size was estimated by comparing with BAC-Tracker™ super-coiled DNA Ladder (Epicentre Bitotechnologies, Madison, WI) after gel-electrophoresis in 0.8% agarose gel. Replicon typing was done by a PCR-based replicon typing (PBRT) method (Carattoli et al., 2005).
Genetic environment of bla CTX-M gene was investigated by PCR and sequencing of the regions surrounding this gene. A combination of IS26 (Eckert et al., 2006), ISCR1 (Brizio et al., 2006) or ISEcp1 (Saladin et al., 2002) forward primers and the CTX-M reverse consensus (MA2) primer (Saladin et al., 2002) were used to investigate regions upstream of the bla genes, and the MA1 primer (Saladin et al., 2002) and reverse primers of IS903, orf477 or mucA genes (Eckert et al., 2006) were used to characterize downstream of the bla genes.
Pulsed-field gel electrophoresis (PFGE) of XbaI (Takara Bio Inc, Shiga, Japan) digested genomic DNA was carried out as described previously (Gautom, 1997). The similarities of the restriction fragment length polymorphisms were analyzed using Bionumerics software, version 4.0 (Applied Maths, Sint-Martens-Latem, Belgium) to produce a dendrogram.
Results
Of 1212 faecal samples examined, 303 (25.0%) showed E. coli growth on CTX-MacConkey agar. Of these 303 E. coli isolates, 121 (39.9%) produced ESBL. Except one isolate from cattle, all of them were swine isolates. These 121 E. coli isolates were subjected to further genotypic characterization. All ESBL-producing isolates were resistant to ampicillin and cephalothin, had cefotaxime MICs that ranged from 4 to >16 μg/mL, and were fully susceptible to imipenem and cefoxitin.
All 121 ESBL-producing E. coli isolates carried bla CTX-M genes belonging to members of CTX-M-1 (n=30) and CTX-M-9 (n=91) families. The most predominant CTX-M–type identified was CTX-M-14 (n=82) followed by CTX-M-15 (n=16). Isolates producing CTX-M-3 (n=9), CTX-M-27 (n=3), CTX-M-55 (n=5), and CTX-M-65 (n=6) were also identified. Among them, single ESBL-producing cattle isolate detected carried bla CTX-M-15 gene. The distribution of 121 bla CTX-M genes according to farms is shown in Table 1. Overall, the bla TEM-1 gene was associated with CTX-M β-lactamases in 55 E. coli isolates carrying bla CTX-M genes. No bla SHV gene was detected among the bla CTX-M-positive isolates.
The transfer of ESBL phenotype was demonstrated from 76 out of 121 bla CTX-M-positive E. coli isolates to the recipient by conjugation experiments (Table 2). PCR analysis showed the presence of respective bla CTX-M genes from all the transconjugants. However, the bla TEM-1 from only 23 out of the 55 E. coli isolates that co-carried bla CTX-M and bla TEM-1 genes got transferred to the recipient E. coli J53. In addition to ESBL phenotype, resistance traits to non-β-lactam antibiotics also co-transferred in many cases along with bla CTX-M genes in the specific plasmid (Table 3).
All E. coli were isolated from pigs, except 08-EM-CF-2 strain, which was isolated from cattle. Etest, Epsilometer test; MIC, minimum inhibitory concentration; ND, not determined; NT, not typable by PCR-based replicon typing; NA, not applicable; CT, cefotaxime; CTL, cefotaxime-clavulanic acid; TZ, ceftazidime, TZL, ceftazidime-clavulanic acid; CHL, chloramphenicol; GEN, gentamicin; NEO, neomycin; STR, streptomycin; SXT, trimethroprim-sulfamethoxazole; TET, tetracycline; —, negative.
Underlined bla genes were transferred to recipient E. coli J53 by conjugation.
Type II was characterized by the presence of ISEcp1 upstream of the bla CTX-M gene but downstream IS903 was absent. Type III showed an identical structure to type II in the upstream region, but mucA gene was found downstream of the bla CTX-M gene.
Plasmid size shown in kilobasepair (Kb).
All E. coli isolates investigated demonstrated at least two plasmids with sizes ranging from 1.5 to 120 Kb (data not shown). However, plasmid DNA isolation from the bla CTX-M-positive transconjugants revealed the presence of the largest plasmid harbored by each donor strain, which ranged in size from 90 to 120 Kb (Table 3). The bla CTX-M-carrying plasmids from E. coli J53 transconjugants were further characterized by PBRT method. Several replicon types, including IncF, IncI1-Iγ, IncFIA, IncFIB, IncFIC, IncHI1, IncX, and IncP, either alone or in combination, were identified in 74 out of 76 transconjugants. The replicon of plasmids from the two transconjugants could not be typed by the PBRT method.
PCR identified ISEcp1 upstream of bla CTX-M genes in 118 out of 121 E. coli isolates (Table 2) and in their 75 corresponding transconjugants. These 118 isolates were classified into three different types based on their genetic environment: type I (n=89), type II (n=26), and type III (n=3). Type I was characterized by the presence of ISEcp1 and IS903 elements upstream and downstream of the bla CTX-M gene, respectively. Type II showed identical genetic environment to type I in the upstream region of bla CTX-M gene, but the downstream IS903 was absent. Type III showed an identical structure to type I in the upstream region, but mucA gene was found downstream of the bla CTX-M gene.
PFGE analysis of a subset of 42 E. coli strains carrying bla CTX-M-14 or bla CTX-M-15 genes from four different farms in which the isolation rate of E. coli strains producing CTX-M β-lactamase was highest demonstrated at least four arbitrary clusters of PFGE profiles (Fig. 1). Whereas, the E. coli strains carrying bla CTX-M-15 genes revealed great genomic diversity, the bla CTX-M-14-positive strains were genetically more homogenous.
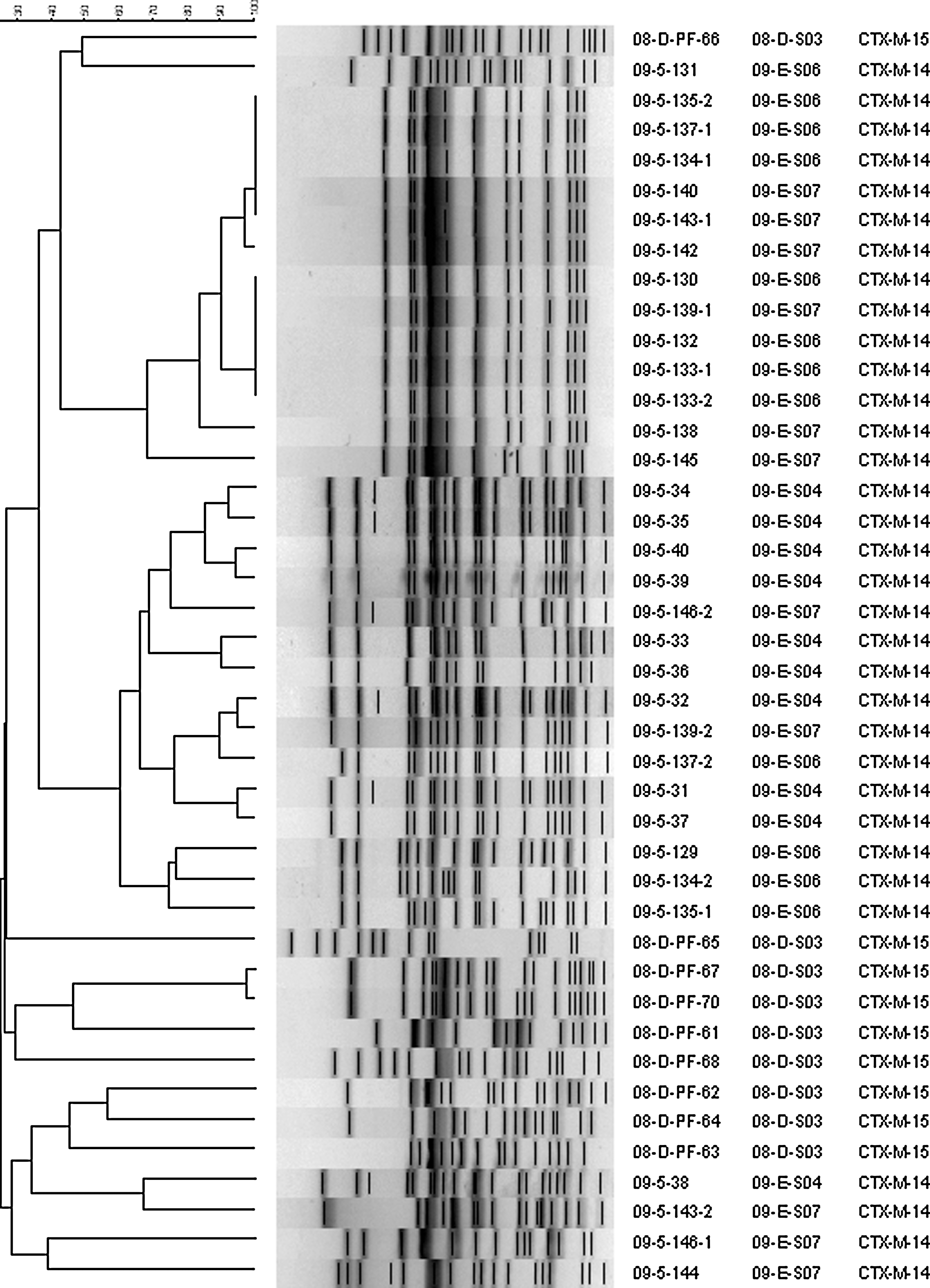
Dendrogram generated by Bionumerics software showing the cluster analysis of XbaI- pulsed-field gel electrophoresis patterns of bla CTX-M-positive Escherichia coli strains isolated from healthy pigs in Korea. Similarity analysis was performed by using the Dice coefficient, and clustering was done by the unweighted-pair group method with arithmetic averages (UGPMA). Details given include the strain, farm number of host animals, and bla CTX-M gene carried by each strain.
Discussion
This study provides the first extensive molecular characteristics of CTX-M–type extended spectrum β-lactamases in E. coli from healthy pigs and cattle in Korea. E. coli from 21.5% (120/558) of the swine fecal samples examined demonstrated CTX-M β-lactamase–production compared to that from cattle fecal samples (1/654, 0.2%). In contrast, higher (6%) prevalence of CTX-M β-lactamase–producing E. coli was reported in fecal samples from cattle from the United States (Wittum et al., 2010). In addition, a high prevalence of E. coli carrying bla CTX-M genes has been previously recovered from healthy pigs and cattle from Hong Kong (Ho et al., 2011). The lower prevalence rate of CTX-M β-lactamase–producing E. coli in healthy animals in Korea may be associated with lower amount of consumption of ceftiofur and other cephalosporins among animals in Korea (NVRQS, 2011). It has been assumed that ceftiofur use in animals may provide selective pressure for the dissemination of the bla CTX-M genes (Tian et al., 2009). Furthermore, all the 121 isolates produced CTX-M β-lactamases belonging to members of CTX-M-1 and CTX-M-9 families. Consistent to our results, dissemination of E. coli harboring members of CTX-M-1 or CTX-M-9 families has repeatedly been reported in Enterobacteriaceae from food animals from Hong Kong (Duan et al., 2006; Ho et al., 2011), China (Tian et al., 2009), and Europe (Blanc et al., 2006; Meunier et al., 2006; Madec et al., 2008). Moreover, recent reports indicated that the significant increase in the incidence of ESBLs in human clinical isolates of E. coli in Korea was due to the dissemination of CTX-M β-lactamases (Song et al., 2009).
In this study, a total of six bla CTX-M alleles (CTX-M-3, −14, −15, −27, −55, and −65) were detected. Except bla CTX-M-55, the bla CTX-M genes characterized in this work were already observed in clinical isolates in human medicine in Korea (Kim et al., 2005; Lee et al., 2009; Tamang et al., 2008). We also detected CTX-M-14 and CTX-M-15 in nontyphoid Salmonella with reduced susceptibility to ciprofloxacin among food animals and humans in Korea (Tamang et al., 2011). In addition, all the CTX-M genotypes observed in this study were recently reported in E. coli isolated from stray dogs in Korea (Tamang et al., 2012), in E. coli from healthy pigs and cattle from Hong Kong (Ho et al., 2011), and in E. coli from pets from China (Sun et al., 2010), which indicate the wide spread of these genes in animals in this region. Of particular interest, in this study, is the predominance and relatively high prevalence of CTX-M-14 (82/558, 14.7%) among the swine E. coli isolates, which is consistent with other reports done earlier among human clinical isolates in Korea in which the most predominant CTX-M–type identified in E. coli was CTX-M-14 (Lee et al., 2009). Similar findings have also been reported from previous investigations on animal isolates from Hong Kong (Ho et al., 2011) and mainland China (Sun et al., 2010) from this region. In contrast, 73% of the strains from pig farms had CTX-M-1, and none of them had CTX-M-9 family enzyme in a study done in Spain (Blanc et al., 2006). The reason for this difference is unknown but it could be interesting to study why CTX-M-14 is predominant in E. coli population both in humans and animals in Asia.
To date, the only report of CTX-M β-lactamases in E. coli from food animals in Korea has been the identification of bla CTX-M-15 in an E. coli isolate from a sick pig and bla CTX-M-14 in E. coli isolates from two sick cattle by our group (Lim et al., 2009). Compared to our previous findings, the prevalence of bla CTX-M genes in swine E. coli has increased several folds from 0.4% (1/249) to 21.5% (120/558) in this study. Similarly, a high number (120/558, 21.5%) of swine fecal samples showed the presence of CTX-M β-lactamase–producing E. coli compared to cattle samples (1/654, 0.2%). Since CTX-M genes are usually located in multidrug resistant plasmids, co-selection of CTX-M genes owing to increasing use of florfenicol, a veterinary chloramphenicol and/or sulfamethazine in pigs for prophylaxis in Korea (NVRQS, 2011) could possibly explain the high prevalence of bla CTX-M genes in swine E. coli isolates, a large majority of which showed resistance to non-β-lactams such as chloramphenicol, tetracycline, gentamicin, or sulfamethoxazole. This is supported by the fact that larger amount of antibiotics was used in pigs (>50% of total consumption) for prophylaxis compared to that in cattle (<10% of total consumption) in Korea (NVRQS, 2011). Similarly, this study showed greater diversity of CTX-M β-lactamases compared to our previous study (Lim et al., 2009). To our knowledge, this study represents the first report of CTX-M-3, CTX-M-27, CTX-M-55, and CTX-M-65 β-lactamases in bacteria-derived from food animals in Korea. Overall, our results showed the wide dissemination of increasing diversity of bla CTX-M genes among healthy pigs, which constitutes a potential public health threat.
A majority of bla CTX-M genes (76/121, 62.8%) transferred to a recipient E. coli, concomitantly with the ESBL phenotype, by mating experiments. Each transconjugant apparently carried a single large plasmid which is difficult and often inaccurate to size. However, 17 of these plasmids belonged to multiple replicon types. This inconsistency may be, in part, due to loss of plasmids by shearing effects or overflow of smaller plasmids during long electrophoresis process. Nevertheless, most bla CTX-M-14-positive transconjugant plasmids (33/46, 71.7%) carried IncF replicon alone or in combination with repFIA/FIB/FIC or/and IncI1-Iγ replicons, suggesting the horizontal dissemination of bla CTX-M-14 gene by IncF conjugative plasmid. This is in agreement with a recent study which reported the spread of bla CTX-M-14 genes by IncF plasmids among human clinical isolates of E. coli in Korea (Kim et al., 2011). In contrast, most bla CTX-M-15-positive plasmids (6/12, 50%) belonged to IncI1-Iγ replicon indicating the role of IncI1-Iγ in the horizontal spread of bla CTX-M-15 genes. In fact, the dissemination of bla CTX-M-15 genes by IncI1-Iγ plasmids in E. coli has been previously documented (Carattoli, 2009). Further, detection of multiple replicons in 17 out of 76 transconjugant plasmids suggests that these plasmids are continuously evolving through replicon sequence recombination, mosaicism, and replicon co-integration or excision events (Osborn et al., 2000).
Detection of ISEcp1 upstream of bla CTX-M genes in more than 97% (118/121) of the E. coli isolates and in their corresponding transconjugants suggests that ISEcp1 may have played a critical role in the efficient capture, expression, and continuous mobilization of bla CTX-M genes, irrespective of CTX-M sequence type. Indeed, ISEcp1 has been frequently found upstream of several CTX-M genes belonging to the CTX-M-1, CTX-M-2, CTX-M-9, and CTX-M-25 families, providing promoter sequences for high-level expression of CTX-M enzymes (Poirel et al., 2003; Saladin et al., 2002). In contrast, PCR amplification of IS26 or ISCR1 upstream of the bla CTX-M genes were negative suggesting that none of these elements contributed to the capture and mobilization of CTX-M genes, as has been previously reported (Meunier et al., 2006). Furthermore, IS903 was detected downstream of bla CTX-M genes belonging to CTX-M-9 family in all 91 E. coli isolates. No orf477 were found downstream of the bla CTX-M genes in the studied isolates. Nevertheless, IS903 and orf477 have been repeatedly reported downstream of most CTX-M-9 and CTX-M-1 family genes, respectively, in several Enterobacteriaceae isolates worldwide (Eckert et al., 2006; Kim et al., 2011).
Our PFGE and plasmid analysis results imply that a combination of clonal expansion and horizontal transmission is spreading bla CTX-M genes among swine E. coli strains in Korea. The clonal spread, accounting for much of the bla CTX-M-14 gene dissemination, occurred both within and between farms, for instance within Farm 09-E-S06 and Farm 09-E-S07 or between them. In contrast, the spread of bla CTX-M-15 genes was attributed to horizontal transmission.
In conclusion, this study represents the first report of CTX-M-3, CTX-M-27, CTX-M-55, and CTX-M-65 β-lactamases in bacterial isolates from food animals in Korea. Our results revealed that the CTX-M β-lactamase–producing E. coli are widely disseminated among healthy pigs but very rare in cattle. Increasing prevalence of bla CTX-M genes in intestinal E. coli of healthy food-producing animals is a matter of concern and should be carefully monitored.
Footnotes
Acknowledgments
This work was supported by a grant from the Animal, Plant, and Fisheries Quarantine and Inspection Agency, Ministry of Food, Agriculture, Forestry and Fisheries, Republic of Korea.
Disclosure Statement
No competing financial interests exist.