
Abstract
Reducing Salmonella in poultry has been a paramount goal of the poultry industry in order to improve food safety. Inclusion of high-fiber fermentable feedstuffs in chicken diets has been shown to reduce the incidence of Salmonella colonization in laying hens, but no work has been performed in growing birds. Therefore, the present study was designed to quantify differences in artificially inoculated cecal Salmonella Typhimurium populations in growing layer chicks (n=60 in each of two replications) fed 0%, 25%, and 50% of their diet (w/w) replaced with alfalfa meal from day (d) 7 to d14 after hatch. Alfalfa supplementation reduced cecal populations of Salmonella by 0.95 and 1.25 log10 colony-forming unit per gram in the 25% and 50% alfalfa groups compared to controls. Alfalfa feeding reduced (p<0.05) the number of cecal- and crop-positive birds compared to controls. Increasing levels of alfalfa increased (p<0.05) total volatile fatty acids (VFA) and the proportion of acetate in the cecum. Surprisingly, alfalfa inclusion did not negatively impact average daily gain (ADG) in birds over the 7-d feeding period. Alfalfa inclusion at 50% of the diet increased (p<0.05) the number of bacterial genera detected in the cecum compared to controls, and also altered proportions of the microbial population by reducing Ruminococcus and increasing Clostridia populations. Results support the idea that providing a fermentable substrate can increase gastrointestinal VFA production and bacterial diversity which in turn can reduce colonization by Salmonella via natural competitive barriers. However, further studies are obviously needed to more fully understand the impact of changes made in diet or management procedures on poultry production.
Introduction
S
High-grain diets are fed to chickens to maximize growth and efficiency, which has allowed the poultry industry to provide a low-cost product to consumers. Altering the diet by including high levels of insoluble fiber could affect growth efficiency and, ultimately, producer profitability (Low, 1993). Diet changes can significantly impact the composition of microbial population of poultry, including populations of foodborne pathogenic bacteria (Corrier et al., 1991; Walker and Duffy, 1998; Woodward et al., 2005). The use of high-fiber diets such as alfalfa meal instead of starvation during forced molting has been shown to decrease cecal Salmonella populations in laying hens (Dunkley et al., 2007a; McReynolds et al., 2006; Woodward et al., 2005), but the effect of alfalfa supplementation on growing chicks has not been examined. Therefore, the present study was designed to quantify differences in artificially inoculated cecal Salmonella Typhimurium populations in growing layer chicks fed 0%, 25%, and 50% of their diet (w/w) replaced with alfalfa meal. Furthermore, we analyzed the effect of these treatments on the cecal microbial population using a bacterial tag diversity amplification method, bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). We hypothesized that the cecal populations of Salmonella would be reduced by increasing levels of alfalfa inclusion, but that growth of the chicks would be negatively impacted by inclusion of this high-fiber feedstuff.
Materials and Methods
Bacterial cultures
Salmonella enterica Typhimurium originally isolated from chickens and maintained in the Food and Feed Safety Research Unit (FFSRU) culture collection was repeatedly grown by 10% (vol/vol) transfer in anoxic (85% N2, 10% CO2, 5% H2 atmosphere) tryptic soy broth (TSB) medium at 37°C. This strain was selected for resistance to novobiocin and nalidixic acid (20 and 25 μg/mL, respectively) by repeated transfer and selection in the presence of sub-lethal concentrations of each antibiotic. This resistant phenotype was stable through multiple unselected transfers in batch culture and through repeated culture vessel turnovers in continuous culture (data not shown). Overnight cultures were harvested by centrifugation (7,500×g, 10 min) and cell pellets were re-suspended in TSB medium. Populations of Salmonella Typhimurium in these cell suspensions were determined to be approximately 2×109 colony-forming units (CFU)/mL, by serial dilution and plating.
Chickens, diet, and experimental design
All procedures in this study were approved by the Institutional Animal Care and Use Committee (IACUC protocol 10-003). Single Comb White Leghorn chicks (SCWL; in two replicates of n=60 birds each) at 1 day (d) of age, were obtained from a local commercial source. Upon hatch, birds were group housed prior to the initiation of this study for an initial growth period of 7 d. Chicks were given free access to water and an unmedicated corn-soybean meal-based layer diet that exceeded National Research Council recommendations (NRC, 1994) that has been previously used in our studies (Dunkley et al., 2007a). Briefly, the diet had the following composition: crude protein, 26%; crude fat, 6%, crude fiber, 4%; lysine, 1.5%; methionine, 1.5%; calcium, 1.1%; and phosphorous, 0.8%. Chickens were housed in an indoor facility with constant artificial light and room temperature was constant at 34°C. Appropriate rearing conditions were provided, as a 0% mortality rate was observed to all the treatment groups involved throughout the duration of the experiment.
On d 7 in each replicate, 20 (10 male and 10 female) chicks were assigned to each of three treatment groups: (1) 0% alfalfa meal, (2) 25% (w/w) alfalfa meal, and (3) 50% (w/w) alfalfa meal. The basal diet used was the same diet used in the acclimation period. Chicks in all treatment groups were provided access to water ad libitum. Alfalfa meal that was used in this study contained >27% crude fiber and 17% crude protein, and had a low metabolizable energy (ME; 1,200 kcal/kg). Chicks were weighed daily individually to obtain treatment group growth data. Dietary treatments were administered for 7 d to determine effects of diet on growth. On d 11, each bird was orally gavaged with 1.2×105 CFU of Salmonella Typhimurium as described above in a 0.5-mL dose.
On d 14, all chicks were euthanized, and the ceca were excised aseptically upon necropsy and were squeezed into sterile tubes (1 g) for dilution (quantitative enumeration) as well as into tetrathionate tubes for qualitative enrichment. Samples from all birds were pooled, stored on wet ice, and shipped overnight for analysis by bTEFAP to Pathogen and Research Testing (Lubbock, TX) and to Lincoln, Nebraska, for volatile fatty acid (VFA) analysis.
Bacterial enumeration
Ruminal, cecal, and rectal contents (10–20 g), as well as excreted feces were serially diluted (1 mL in 10-fold increments) in phosphate-buffered saline (PBS; pH 6.8) and directly plated on Brilliant Green Agar supplemented with novobiocin (20 μg/mL) and nalidixic acid (25 μg/mL; BGANN) plates. Colonies that grew on agar plates after 24 h of incubation were directly counted (quantitative enumeration with a lower limit of detection of 101 CFU/mL). To qualitatively confirm the presence of inoculated Salmonella Typhimurium, intestinal contents and epithelial tissue samples as well as feces were enriched in tetrathionate broth at 39°C for 24 h followed by a secondary enrichment in Rappaport-Vasilladis broth at 42°C for 24 h with final plating on BGANN plates. Plates that contained typical Salmonella colonies after 24 h incubation were classified as positive for experimentally introduced Salmonella Typhimurium. Random colonies were picked during the course of the study and examined via growth on triple sugar iron agar (TSI) and lysine iron agar (LIA) slants that were incubated at 37°C for 18 h to verify that the colonies growing on the BGANN plates were indeed Salmonella Typhimurium.
The bTEFAP analysis
The bTEFAP was performed, which is based upon a partial ribosomal amplification followed by pyrosequencing (PRAPS) (Dowd et al., 2008). Full technical details of the bTEFAP procedure used are provided in previous studies, and are omitted here for brevity (Bailey et al., 2010; Callaway et al., 2010; Callaway et al., 2009). Total genomic DNA was extracted from fecal samples using a QIAamp stool DNA mini kit and its manufacturer's recommended methods (Qiagen, Valencia, CA). DNA samples were quantified using a Nanodrop spectrophotometer (Nyxor Biotech, Paris, France).
VFA analysis
Each cecal sample was diluted with an equal weight of distilled deionized water, shaken for 30 s, and centrifuged at 10,000 x g for 2 min to separate solids from the liquid portion of the sample. A 500-μL volume of the liquid extract was then acidified by adding 50 μL of 3 M HCl, mixed well, and centrifuged again for 2 min to separate additional acid-precipitated solids from the liquid extract. Alcohols (ethanol, propanol, isobutanol, butanol, and pentanol), VFA (acetate, propionate, isobutyrate, butyrate, isovalerate, valerate, isocaproate, caproate, heptanoate, and caprylate), and aromatic compounds (phenol, ρ-cresol, 4-ethyl phenol, indole, skatole, benzoate, phenylacetate, and phenylpropionate) were immediately quantified using a Agilent 7890 gas chromatograph (Agilent Technologies, Santa Clara, CA) equipped with a flame ionization detectors. A 1-μL volume was introduced by autoinjector into a split/splitless inlet operated at 275°C and at a 10:1 split. Individual compounds were then separated on a DB-Wax (Agilent Technologies) column (30 m length, 0.25 mm diameter, 0.25 mm film thickness) using a ramped-flow (0.25 mL/min for 0.6 min; 20 mL/min to 3.5 mL/min for 0.4 min; 20 mL/min to 0.25 mL/min for 0.5 min; 20 mL/min to 3.5 mL/min) and ramped-temperature (50°C for 2 min; 30°C/min to 170°C; 15°C/min to 230°C; 5°C/min to 240°C) program, and quantified using standard solutions of alcohols, VFAs, and aromatic compounds treated identically to the cecal samples.
Reagents and supplies
Unless otherwise noted, all media and agar were purchased from Difco Laboratories (Sparks, MD). Reagents and antibiotics were obtained from Sigma Chemical Co. (St. Louis, MO).
Statistical analysis
Salmonella Typhimurium CFU/g were log10 transformed. Treatment groups were compared at each time point by the Mixed procedure of SAS (SAS Institute Inc., Cary, NC). Time by treatment interactions were discounted due to the natural decay of Salmonella Typhimurium populations in this artificially inoculated model; therefore, only within-time comparisons were performed. Numbers of Salmonella-positive birds were compared with Fisher's Exact test. Significance was determined at p<0.05.
Results
Inoculated Salmonella populations in the cecal contents of chickens fed 25% and 50% alfalfa meal were reduced compared to controls by 0.95 and 1.25 log10 CFU/g (Fig. 1), the decrease was only reduced (p<0.08) at the 50% level due to variance of shedding and the relatively low initial dose of Salmonella. The number of birds that were positive for Salmonella Typhimurium in the crop were lower (p<0.01) in the 25% and 50% alfalfa groups than in controls, but these groups were not different from each other (Fig. 2). Birds that were fed 25% or 50% alfalfa had fewer birds that were positive for Salmonella in the cecal contents than did chicks in the control group (p<0.05), but there was no difference in the number of positive birds between the 25% or 50% alfalfa groups (p>0.2; Fig. 2).
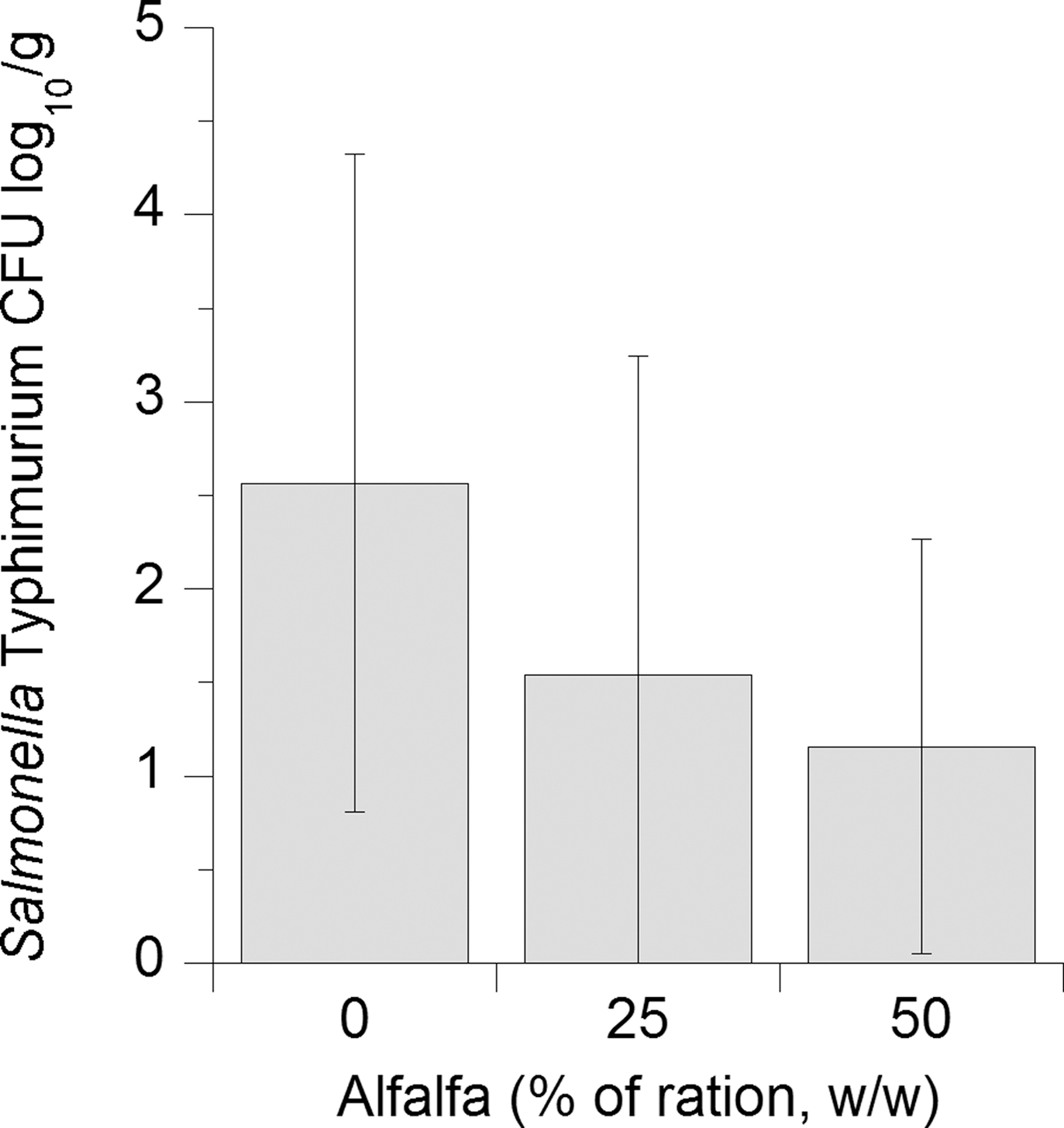
Cecal populations (CFU/g feces, log10) of Salmonella Typhimurium in chicks (n=40/treatment) fed 0%, 25%, or 50% alfalfa meal. Error bars indicate standard deviations.
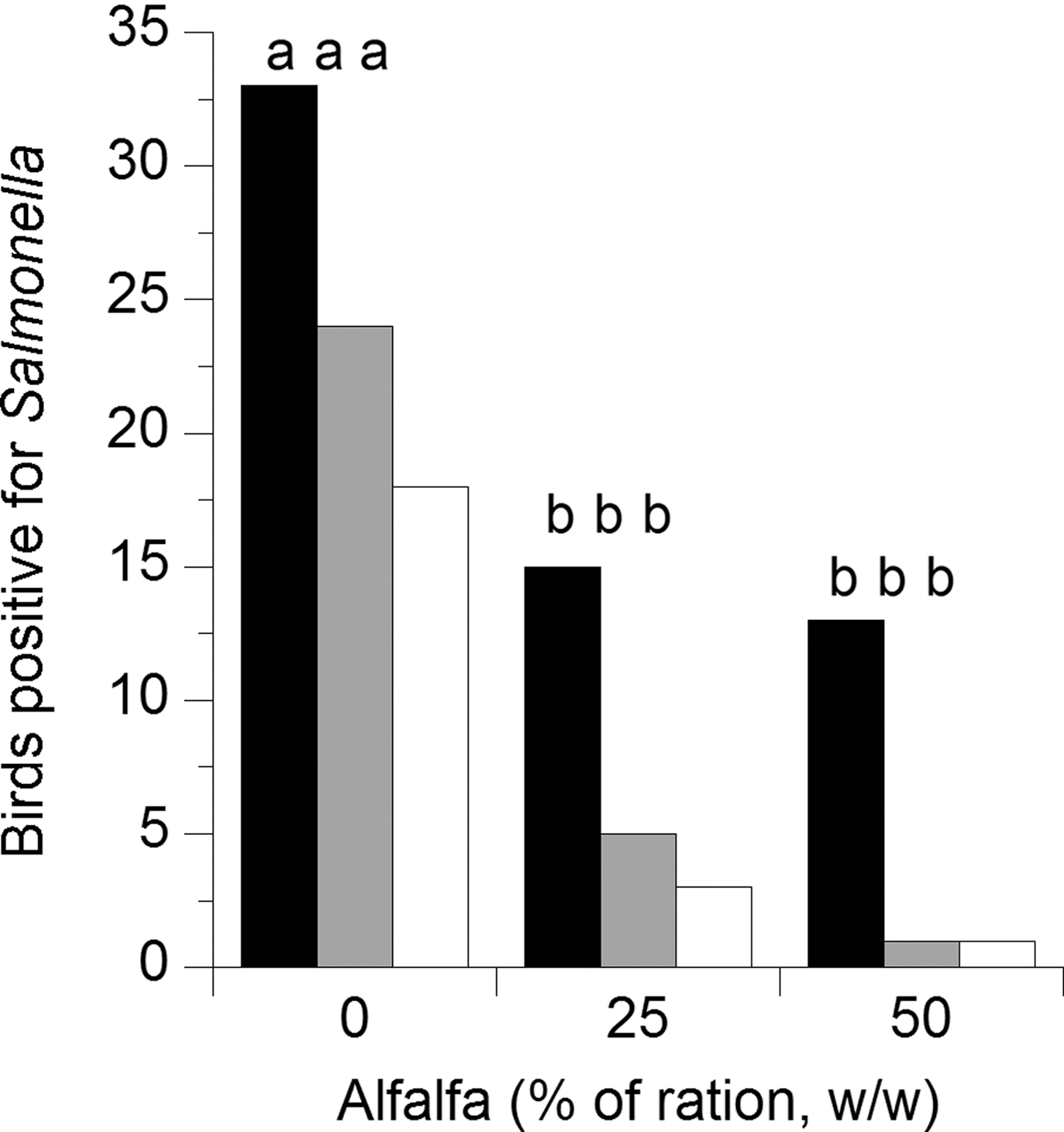
Number of chicks (n=40/treatment) positive for Salmonella Typhimurium after enrichment. Birds were fed 0%, 20%, or 50% alfalfa meal. Gray bars, number of birds positive in the crop; black bars, number of birds with Salmonella-positive cecal contents; white bars, number of birds with Salmonella in both the crop and cecum. Bars with different superscripts differ from control by p<0.05.
VFA concentrations in the cecal contents were affected by dietary inclusion of alfalfa (Fig. 3). Total VFA concentrations were increased (p<0.05) compared to controls in the 25% and 50% alfalfa-fed groups. This increase was attributed to an increase (p<0.05) in both acetate and butyrate concentrations in both the 25% and 50% groups, while propionate concentrations were only increased (p<0.05) in the 50% alfalfa group. Concentrations of all VFA were highest (p<0.05) in the ceca of birds fed 50% alfalfa diets.
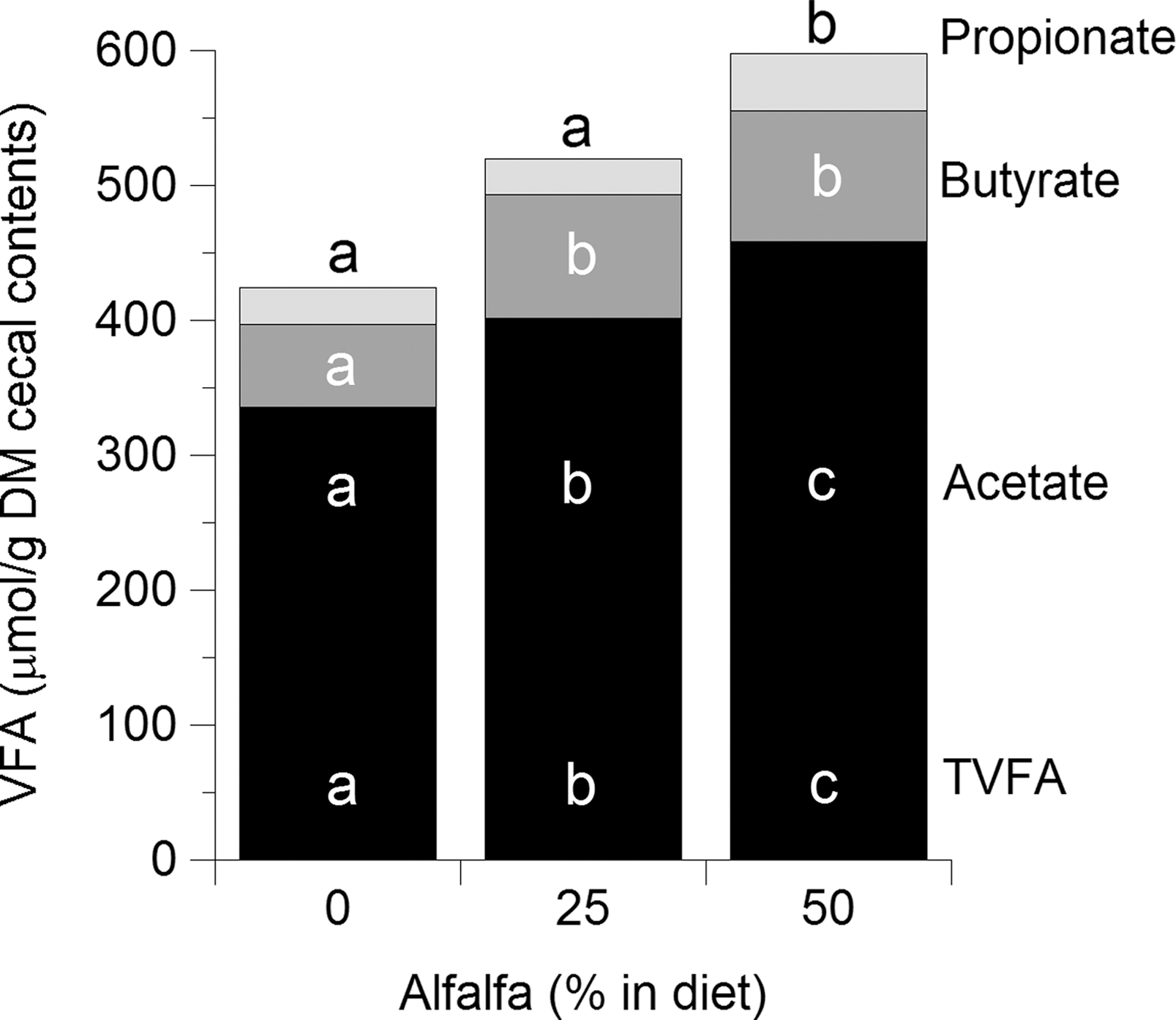
Volatile fatty acid (VFA) concentrations in cecal fluid (μmol/g dry matter [DM] cecal contents) from chicks fed 0%, 20%, or 50% alfalfa meal in their diets. Concentrations of acetate, propionate, butyrate, and total VFA are presented. Bars with different superscripts differ by p<0.05.
The average daily gains (ADGs) for the 7-d period following initiation of the 25% and 50% alfalfa feeding are shown in Figure 4. No differences were detected between the ADG of any of the three treatments. Body weight was numerically higher in the 25% and 50% alfalfa groups at the conclusion of the study (132 and 125 g/bird, respectively), but this was not different from the controls (119 g/bird).

Average daily gain (ADG; g/bird/day) of chicks fed 0%, 20%, or 50% alfalfa meal in their diets. ADG is presented as mean over 7 days from initiation of alfalfa inclusion in the diet. Error bars indicate standard deviations.
Bacteria were detected in all cecal samples across treatments and pyrosequencing results indicated some differences between treatment groups based on the amount of alfalfa in the diet (Table 1). The effect of alfalfa feeding on cecal bacterial diversity is reflected in the increasing numbers of genera and species detected in the ceca of birds fed 25 and 50% alfalfa diets compared to controls (Table 1). The composition of the microbial ecosystem changed. Ruminococcus proportions were reduced by 40–50% in the ceca of birds fed 25% and 50% alfalfa compared to controls (Table 1). Conversely, proportions of Clostridium increased twofold (p<0.06), and cecal Escherichia increased up to sevenfold by 25% and 50% alfalfa feeding. Cecal proportions of Eubacterium, Oscillospira, Klebsiella, and Bacteroides were all increased at least twofold by alfalfa feeding (Table 1 and Fig. 5). Interestingly, populations of Shigella were detected by pyrosequencing at very low levels (but were not tested for via cultural methods) in all diets and were increased numerically in the ceca of chickens fed 25% and 50% alfalfa meal, but this must be taken with a note of caution as Shigella was not cultured.
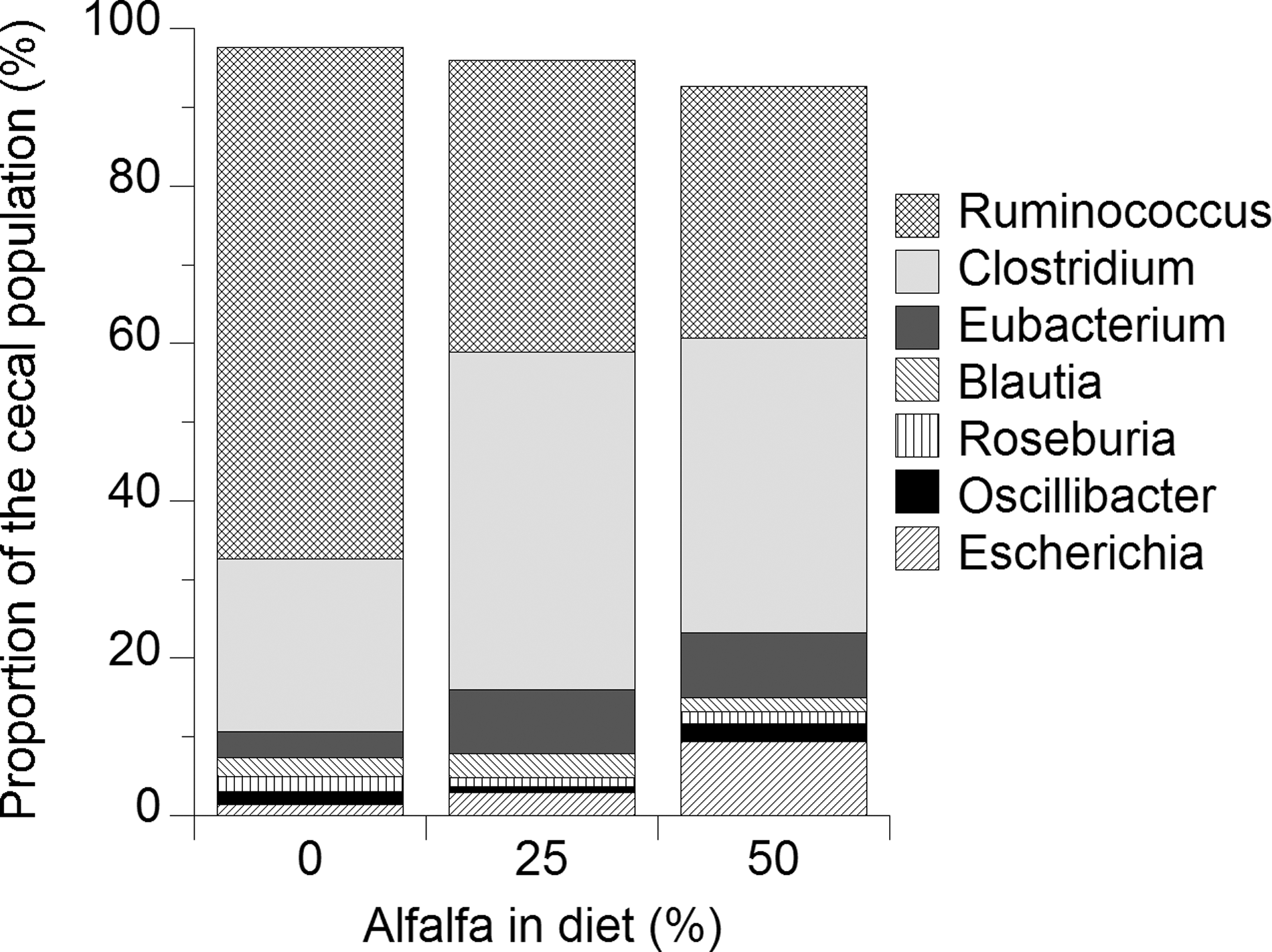
Proportions of cecal bacterial populations (%) from chicks fed 0%, 20%, or 50% alfalfa meal in their diets. The seven most prevalent bacterial genera from Table 1 are depicted.
Rows with different superscripts differ by p<0.06.
SD, standard deviation.
Discussion
Salmonella of many serotypes can colonize poultry, and is most frequently isolated from the crop and the ceca of chickens (Jorgensen et al., 2002; Linton et al., 1985), but can also penetrate tissues and be transmitted via eggs (Braden, 2006). The majority of human foodborne Salmonellosis cases are linked to poultry consumption (Guo et al., 2011). Therefore, in order to most dramatically improve human food safety and health, it is critical to understand how Salmonella survives and thrives in the chicken so that amplification and transmission cycles can be interrupted (Callaway et al., 2007; Vandeplas et al., 2010).
Several preharvest Salmonella reduction strategies have been proposed in poultry (Vandeplas et al., 2010), and some of these have involved dietary inclusions or changes (Corrier et al., 1997). Feeding of high-fiber dietary components to molting hens has been shown to decrease cecal Salmonella populations compared to starvation (Dunkley et al., 2007a; McReynolds et al., 2006; Woodward et al., 2005). This anti-Salmonella effect of alfalfa feeding was observed in previous studies both with Salmonella Enteriditis and Typhimurium serotypes, and is thought to be related to the maintenance of a more “normal” and diverse bacterial population in the gastrointestinal tract (Callaway et al., 2009; Dunkley et al., 2007b).
Alfalfa has a relatively high fiber content and has a long transit time in the gastrointestinal tract of poultry, resulting in the cecal bacterial fermentation of alfalfa that produces VFA. VFAs have been shown to be toxic to bacteria (Wolin, 1969) and have been used as mechanisms for reducing pathogen populations in the gut (Prohaszka and Baron, 1983). In the present study, alfalfa feeding increased both total VFA as well as proportions of acetate and butyrate in the cecum. Correlating the VFA concentrations with the cecal Salmonella populations and colonization results suggests that the production of VFA in the cecum, at least in part, reduced Salmonella populations. Furthermore, VFA production from alfalfa fermentation in the crop, while not measured in this study, is hypothesized to play a role in reducing the number of observed Salmonella positive crops observed.
Chickens are fed high-grain diets in order to maximize growth and efficiency (NRC, 1994), which has allowed the poultry industry to provide a rapid, consistent, wholesome product to consumers. Altering this diet by including high levels of insoluble fiber could potentially affect growth efficiency and ultimately, producer profitability (Low, 1993). Therefore, we hypothesized that the inclusion of 25% and 50% alfalfa meal in the diet would negatively impact growth of the birds in the present study. Surprisingly, however, ADG of the chicks was not affected by alfalfa inclusion during the short study duration. However, because newly hatched chicks are most susceptible to Salmonella colonization (Snoeyenbos et al., 1978), this result suggests that the dietary inclusion of alfalfa meal might be a method to reduce Salmonella colonization at least during that initial susceptible period, without significant negative impact on growth rate. However, more studies are needed to determine the long-term effects on growth and Salmonella status of birds fed alfalfa during the early grow-out phase.
The microbial population of the gastrointestinal tract can profoundly alter the health and productivity of the host animal (Lyte, 2010). Microbes in the gastrointestinal tract interact with each other as well as their host animal, and many of these multifactorial interactions are undefined because of a lack of knowledge about the composition of the microbial population of the gastrointestinal tract (Lyte, 2010). Dietary changes have been shown to have widespread impacts on the microbial ecosystem of the gut, and these changes can impact animal health, productivity and food safety (Awad et al., 2011; Jacob et al., 2009). The development of molecular methodologies have allowed researchers to utilize rapid sequencing technologies such as pyrosequencing to quantitatively evaluate the microbial diversity of the gut in novel ways (Roesch et al., 2007).
Molecular-based research has suggested that the chicken gastrointestinal microflora contains more than 650 species of which more than half are from previously unknown bacterial genera (Apajalahti et al., 2004). In the present study, only 92 different genera were isolated from all the chicken ceca, which is fewer than previous studies have found when examining molting layer hens (Callaway et al., 2009). However, this result was unsurprising because the mature gastrointestinal population of laying hens is more complex than the population found in the young growing birds utilized in the present study. Alfalfa supplementation increased the number and diversity of genera isolated from the ceca of birds, perhaps due to the increased availability of fermentable substrate. Ruminococcus was the most prevalent bacterial genus in the cecal contents of birds fed the control diet, but when alfalfa was included in the diet Clostridium became more prevalent than Ruminococcus (Table 1). This shift in Ruminococcus with a concomitant increase in Clostridia is similar to that seen when alfalfa was included in molting rations compared to starvation (Callaway et al., 2009); however, in the present study, the proportions of these genera are much (>10-fold) higher. Interestingly, Ruminococcus is best-known as a ruminal bacteria involved in fiber degradation (Helaszek and White, 1991), but in the present study populations were decreased in chicken ceca with increasing levels of fiber, but did not appear to be related to cecal pH (data not shown). We hypothesize that this apparent contradiction can be attributed to the resident time of feedstuffs in the chicken cecum being too short to allow Ruminococcus populations to exploit the fiber fermentation niche. Clostridia have been implicated in animal health conditions, such as necrotic enteritis (Hofacre et al., 2003; Miller et al., 2010); however, in the present study, these effects were not detected, but that would clearly need to be investigated prior to widespread introduction of alfalfa feeding to reduce Salmonella in broilers. Overall, large changes in the bacterial composition of the gastrointestinal tract in this study were limited primarily to a few genera likely due to the relatively low microbial diversity of the young birds.
Conclusion
Salmonella is a threat to the safety and wholesomeness of poultry products, and methods to reduce the incidence of this foodborne pathogen are of great importance to improving food safety. Preharvest strategies to reduce the prevalence and concentrations of these pathogenic bacteria have been developed, but few to date have been widely implemented. Newly hatched chicks are susceptible to Salmonella colonization; therefore, methods to reduce the penetration of Salmonella at this vulnerable stage are critical. Supplementing diets with 25% or 50% alfalfa meal reduced cecal populations of Salmonella by approximately 10-fold, and reduced the numbers of Salmonella-positive birds in the crop and cecum but did not reduce the ADG of the chicks. While feeding chickens 50% alfalfa throughout their growth period is not feasible financially or logistically, these results support the idea that providing a fermentable substrate to the gastrointestinal population can increase VFA production and gastrointestinal bacterial diversity which in turn has the potential to reduce colonization by Salmonella via natural competitive barriers. However, further studies are obviously needed so that we can more fully understand the impact of changes made in diet or management procedures of poultry production.
Footnotes
Acknowledgments
We would like to thank Dr. Lisa Durso for her critical evaluation of the data and manuscript.
Disclosure Statement
No competing financial interests exist.