
Abstract
Throughout Southeast Asia and China, eating raw and or partially cooked cyprinid fish causes liver (hepatobiliary) disease and cancer (cholangiocarcinoma) due to fishborne zoonotic trematodes (FZT), in particular Clonorchis sinensis and Opisthorchis viverrini. The primary source of transmission is by native fish, but aquaculture fish are also reported to have high infective potential. Here, a cross-sectional survey of FZT in fish farms was conducted in an endemic area in Khon Kaen Province, Thailand. By using conventional and polymerase chain reaction (PCR) methods, we detected O. viverrini and FZT metacercariae (Centrocestus formosanus and Haplorchis taichui) in two popular fish species, Barbonymus gonionotus (silver barb) and Cirrhinus mrigala (mrigal), from aquaculture farms. Both species were infected in five of six farms examined by PCR but not by conventional methods, yet the prevalence of FZT metacercariae in aquaculture fish was high (46.9%). In addition to O. viverrini (17.1%), the native fish Cyclocheilichthys armatus and Hampala dispar had a prevalence of FZT of 81.4%, which included 5.7% for C. formosanus and 17.1% for H. taichui by conventional method. To our knowledge, this is the first discovery of O. viverrini in aquaculture fish in Thailand. More comprehensive studies are required to determine if human-induced disease transmission coupled with natural transmission cycle occurs throughout the aquaculture industry in the region. This has significant impact on food quality and safety, and provides the basis for the development of an effective strategy for the prevention and control of foodborne diseases.
Introduction
Since native fish stocks are inadequate to supply consumers with their needs, there is a growing demand for aquaculture fishery products (FAO, 2010). To accommodate this demand, there has been an increase in the aquaculture industry, especially in the endemic regions of FZT in Asia, which supplies 90% of aquaculture fish worldwide (FAO, 2010). Aquaculture in farms and natural water sources located in endemic areas are at risk of being contaminated by local parasites, which can then be dispersed through the market chain (Phan et al., 2010b).
The presence of FZT in aquaculture fish has been reported in southern China (Fang, 1994; Lun et al., 2005) and Vietnam. Among these FZT, C. sinensis metacercariae were also detected but with a low prevalence (Phan et al., 2010a; Phan et al., 2010b). In Lao People's Democratic Republic (PDR), native fish were also reported to harbor several types of trematode metacercariae, including O. viverrini, in the Num Ngum reservoir and other areas in Vientiane (Ditrich et al., 1990) and in Khammouane (Kobayashi et al., 1996, 2000; Vannachone et al., 1998), Champassak (Chai and Hongvanthong, 1998), and Savannakhet provinces (Rim et al., 2008). In Thailand, there have been studies of parasite infections in native fish in many areas, especially in the north and northeast of the country (Kumchoo et al., 2005; Noikong et al., 2011; Wongsawad and Wongsawad, 2011). For example, 13 and 16 species of cyprinids were collected from Chiang Mai and Sakon Nakhon with overall prevalences of metacercarial infection of 84.5% and 47.6%, respectively. To date, only one study has attempted to apply the Hazard Analysis Critical Control Point (HACCP). This was aimed at controlling FZT in a fish farm in Chiang Mai where O. viverrini was not present (Khamboonruang et al., 1997).
The purpose of this study was to determine whether aquaculture fish are infected with FZT metacercariae, including O. viverrini, as determined by conventional methods compared to species-specific polymerase chain reaction (PCR) primers for O. viverrini, C. sinensis, and other FZTs. This article will provide an assessment and analysis of risk factors of foodborne transmission of FZT by aquaculture.
Materials and Methods
Study areas and sample collection
The study was conducted from November 2008 to February 2009. Based on primary screening of aquaculture farms in Mueang and Nong-Rue district (Georeference coordinates: 16°26′36.89″N-16°45′91.34″N; 102°50′26.52″E-102°83′90.69″E), there were eight out of 16 farms that produced and supply cyprinid fish to farmers. All eight farms were recruited for FZT analyses in this study. In these farms, fish fry produced from local hatcheries were reared in fish ponds filled with natural freshwater and were fed with commercial feed pellets (CP Worldwide, Bangkok, Thailand) until they reached fingering size. All aquaculture sites were similar in structure and procedures. Native fish samples originated from Phu Wiang and Mueang districts (Georeference coordinates: 16°24′51.50″N-16°40′46.67″N; 102°53′54.93″E-102°21′40.93″E), Khon Kaen Province, northeast Thailand. The study sites in these three districts in Khon Kaen Province were selected because they are known as endemic areas of the liver fluke (Elkins et al., 1996; Saowakontha et al., 1993).
The species of aquaculture fish examined were Barbonymus gonionotus (silver barb) and Cirrhinus mrigala (mrigal) because they are the most common cyprinid fish species used for aquaculture in this part of Thailand. To provide information on prevalence external to aquaculture sources within the areas chosen, the native cyprinid species Cyclocheilichthys armatus (Sai Tan Ta Khao) and Hampala dispar (Kra Sup Jut) were examined from dams and water courses. These two species were chosen because they are known to be infected with FZT, including O. viverrini (Srisawangwong et al., 1997). Based on the previous records of the prevalence of FZT in native fish (Vichasri et al., 1982; Wongsawad and Wongsawad, 2011) and aquaculture fish (Chi et al., 2008), a sample size of about 15–20 fish was considered to be adequate for this study.
Detection and identification of metacercariae by conventional method
Following the collection of fish from farms or markets, they were placed on ice and transported to the laboratory, where they were placed in cold storage (4°C) until they were examined for metacercariae, which took place within 7 days of collection. The length and weight of each fish were recorded, and metacercariae isolated and recovered using a standard pepsin digestion procedure as previously described (Sithithaworn et al., 1997). We adopted a similar protocol to that used by Phan et al. (2010b) to recover and identify FZT metacercariae with the following modifications: the metacercariae were counted and identified individually by a stereo and a compound microscope (Srisawangwong et al., 1997). After examination, morphologically unidentifiable metacercariae recovered from each species of fish were each pooled and subjected to PCR analyses. Samples of identified metacercariae were also confirmed by PCR.
Identification of metacercariae by PCR
Pools of FZT metacercariae from each species of fish were extracted for DNA using the phenol/chloroform method: 100 μL of distilled water was added and boiled at 95°C for 10 min. Sodium dodecyl sulphate and proteinase K were added to a final concentration of 1% and 0.1 mg/mL, respectively. The homogenate was incubated at 37°C for 4–5 h. The lysate was extracted with phenol followed by ethanol precipitation. The DNA was re-suspended in 10 mM Tris-HCl, 1 mM EDTA, pH 8.3 (TE), and used for PCR amplification (Maniatis et al., 1982).
Amplification of genomic DNA of FZT metacercariae was carried out using the following specific primers. For O. viverrini, primers used were Trem25F (5′ acaggaagtggaaccgtgtc 3′) and OV25-4R (5′ aatgaacggaaatcgtgacc 3′), and the PCR amplicons were approximately 229 bp (Parvathi et al., 2008). PCR assays were carried out in a final volume of 30 μL consisting of PCR buffer (10 mM Tris-HCl [pH 8.4], 50 mM KCl, 2 mM Mg2+), 0.04 mM each deoxynucleoside triphosphate, 0.05 U Taq DNA polymerase (Intron Biotechnology Inc., SeongNam-Si Gyeonggi-do, Korea), and 0.3 pmol each of forward and reverse primers. Thermocycling conditions included an initial denaturation at 95°C for 5 min, 35 cycles of 95°C for 1 min, annealing at 60°C for 1 min, extension at 72°C for 1 min, followed by a final extension at 72°C for 5 min.
Primers used for C. sinensis were CS1F (5′ ccaaccgagttggtcaagtt3′) and CS1R (5′caatccaacgcactctctga3′), which gives the PCR amplicons of approximately 283 bp (Parvathi et al., 2007). PCR assays were carried out in a final volume of 25 μL consisting of PCR buffer (10 mM Tris-HCl [pH 8.4], 50 mM KCl, 2 mM Mg2+), 0.25 mM each deoxynucleoside triphosphate, 0.06 U Taq DNA polymerase (Intron Biotechnology Inc.), and 1 pmol each of forward and reverse primers. Thermocycling conditions included an initial denaturation at 95°C for 3 min, 40 cycles of 95°C for 1 min, annealing at 62°C for 1 min, extension at 72°C for 1 min, followed by a final extension at 72°C for 10 min.
We designed two new specific PCR primers for H. taichui and C. formosanus from internal transcribed spacer 2 sequences (ITS2). Each primer was checked for specificity and had no cross reactivity with O. viverrini or C. sinensis. The H. taichui primer used was LC1 (5′cgagtatcgatgaagaacgcagc 3′) and HT4sp (5′gtgcacaaagaattgcatgg 3′). PCR amplicons were approximately 558 bp, while the PCR primers used for C. formosanus were LC1 (5′ cgagtatcgatgaagaacgcagc 3′) and Centsp (5′ccaatgccgagatcacagacaag 3′). PCR amplicons were approximately 367 bp. PCR assays were carried out in a final volume of 25 μL consisting of PCR buffer (10 mM Tris-HCl [pH 8.4], 50 mM KCl, 2 mM Mg2+), 0.2 mM each deoxynucleoside triphosphate, 0.05 U Taq DNA polymerase (Intron Biotechnology Inc.), and 0.2 pmol each of forward and reverse primers. Thermocycling conditions included an initial denaturation at 94°C for 1 min, 30 cycles of 94°C for 1 min, annealing at 55°C for 1 min, extension at 72°C for 3 min, followed by a final extension at 72°C for 5 min. All the PCR reactions were carried out in a PTC-200 thermal cycler (M.J. Research Inc., Waltham, MA). The amplified PCR products underwent electrophoresis using a 1.5% agarose gel containing 0.1 mg/mL ethidium bromide, and the gel was analyzed using the Gel Documentation System.
Statistical analysis
Data were analyzed by SPSS version 16. Chi-squares tests were used to compare prevalence of FZT between species of fish and the size of fish. Mann-Whitney U-tests were used to compare intensity of metacercarial infection between species of fish.
Results
Prevalence and intensity of FZT metacercariae based on conventional method
A total of 232 aquaculture and native fish were examined individually for FZT metacercariae using standard morphological characters. Aquaculture fish included 113 B. gonionotus (silver barb) from six farms and 49 C. mrigala (mrigal) from two farms in Khon Kaen city district (Mueang), while native fish included 50 C. armatus from Ubonratana dam and Pra Kue stream, with an additional 20 H. dispar from Ubonratana dam (Table 1). The majority of aquaculture fish examined were fingerlings and juveniles. For B. gonionotus, mean length ranged from of 4.9 to 18.6 cm and mean weight ranged from 1.3 to 83.2 g. For C. mrigala, mean length ranged from of 9.4 to 10.9 cm and mean weight ranged from 7.4 to 10.0 g. The mean width of aquaculture fish was similar between species and ranged from 1.4 to 5.8 cm. The native fish examined were larger and heavier than aquaculture fish (Table 1).
Both species of aquaculture fish from all eight farms were infected with non-taxonomically definable FZT metacercariae with an average prevalence of 46.9%. A high prevalence was found in five of eight farms, ranging from 46.7% to 89.5%. The remaining farms had lower prevalences, ranging from 5.0% to 15.8%. Average prevalence of FZT across all farms for C. mrigala was 38.8% and for B. gonionotus 50.4%. This was not significantly different. The mean intensity of FZT infection in C. mrigala was 22.9±51.5 (mean±SD) metacercariae per fish and 6.9±13.6 metacercariae per fish for B. gonionotus, which was also not significantly different.
For morphologically identifiable FZT metacercariae, only C. formosanus were identified in three farms with an average prevalence of 9.2%. The average prevalence of C. formosanus in C. mrigala (n=49) was 20.4% and in B. gonionotus 4.4% (n=113), with that of C. formosanus in C. mrigala being significantly higher than in B. gonionotus (p<0.001). There was also a significantly higher intensity (p<0.001) of C. formosanus infection in C. mrigala (4.6±15.3 metacecariae per fish) compared to B. gonionotus (0.4±2.0 metacercariae per fish).
The two species of native fish were infected with FZT metacercariae with an average prevalence of 81.4% and mean intensity of 47.2 (SD=92.1). A high prevalence of FZT was found in both species, namely, 86% in C. armatus and 70.0% in H. dispar. These were not significantly different. Table 2 shows that the average prevalence of FZT in native fish (47.2±92.1) was significantly higher (p=0.026) than aquaculture fish (11.7±31.2).
Currently, many FZT have not undergone rigorous taxonomic classification to the species level.
FZT, fishborne zoonotic trematodes; P, prevalence; I, intensity.
—, not detected.
The following metacercariae were found in native fish: C. formosanus, H. taichui, and O. viverrini. For all species of fish (n=70), the average prevalence of C. formosanus metacercariae was 5.7%, and 17.1% for H. taichui and O. viverrini. The average prevalence of H. taichui in H. dispar was 25% and C. armatus was 35%, whereas the average prevalence of C. formosanus and O. viverrini in C. armatus was 13.3% and 24%, respectively.
Molecular identification
PCR was employed to identify both conventionally identifiable as well as unidentifiable FZT metacercariae in aquaculture and native fish. The sample size from the six farms was sufficient to conduct PCR analysis (Table 3). This revealed that fish in five of the six farms were infected with FZT metacercariae of known species, whereas conventional methods detected such FZT in only three farms (Table 2). Furthermore, PCR analysis of unidentifiable metacercariae detected two more species of FZT (O. viverrini and H. taichui) compared to conventional methods. Typical examples of the PCR analyses of pooled unidentifiable metacercariae isolated from aquaculture fish samples based on specific primers for O. viverrini, C. formosanus and C. armatus are shown in Figure 1.
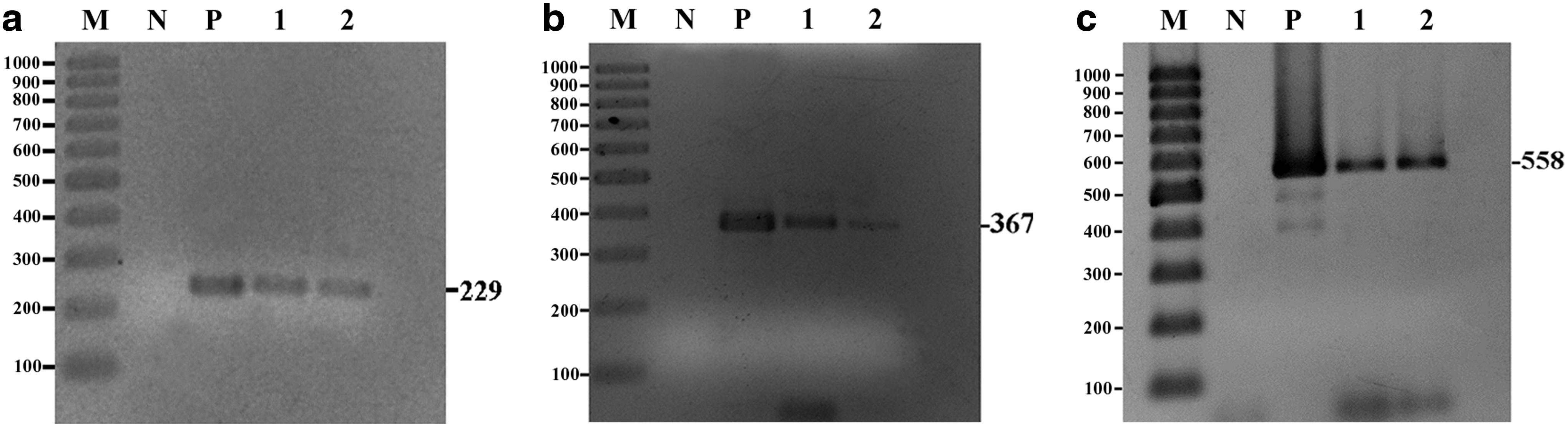
Polymerase chain reaction (PCR) analyses of pooled unidentifiable fishborne zoonotic trematodes (FZT) metacercariae isolated from aquaculture fish samples.
FZT, fishborne zoonotic trematodes; PCR, polymerase chain reaction.
For unidentifiable metacercariae, PCR revealed the presence of O. viverrini in four of six farms in both B. gonionotus and C. mrigala. PCR detected C. formosanus in three of six farms, compared with two of six farms using conventional methods. H. taichui was detected by PCR in two of six farms (Table 3). It was not detected by conventional methods in any of the six farms. For unidentifiable FZT in native fish, PCR detected O. viverrini in H. dispar and C. formosanus in both C. armatus and H. dispar (Table 3).
Discussion
Most studies of FZT to date have used morphological methods to assess trematode infections in fish. The morphological examination of metacercaria from fish is sometimes problematical when encountering atypical metacercariae and is not effective for unidentifiable metacercariae. To compliment this conventional method, for instance, O. viverrini–specific PCRs have been developed and used to detect different life cycle stages of the parasite, including metacercariae in fish (Duenngai et al., 2008; Parvathi et al., 2008; Wongratanacheewin et al., 2001).
The data presented here on native fish show the expected high prevalence and intensities of infection found in previous studies (Sithithaworn et al., 1997) indicating that the probability of a suitable individual host being infected is high. This is consistent with high prevalence levels in five of the eight fish farms. In addition, these results confirm and extend the observations reported by (Chi et al., 2008) that juveniles produced in the nursery aquaculture can be highly infected with FZT. Previous reports from Vietnam have revealed that, in the Mekong Delta, 1.2–29.7% of aquaculture fish were infected with FZT metacercariae (Thien et al., 2009; Thu et al., 2007). In the north of Vietnam, the prevalence of FZT was even higher at 44.6% in Nghe An Province (Chi et al., 2008) and >50% in Nam Dinh Province.
In terms of species diversity, in the present study we found O. viverrini, C. formosanus, and H. taichui in aquaculture fish (silver barb and mrigal). In Vietnam, similar species of FZT (C. formosanus and H. taichui) were also found, with H. pumilio as the predominant trematode species (Thien et al., 2009; Thu et al., 2007). The liver fluke, C. sinensis, was found in silver carp (Hypophthalmichthys molitrix) but with low prevalence (Phan et al., 2010b). Recently C. sinensis has been reported in Thailand (Traub et al., 2009); however, in this study, no C. sinensis were detected in either native or aquaculture fish.
Our analyses of unidentifiable metacerariae by PCR shows that this is a useful technique to support the conventional method in that it revealed the presence of O. viverrini in silver barb and mrigal originating from 50% of the total aquaculture fish farms examined. Water intake in aquaculture fish farms was from a natural reservoir in an endemic area which flows into ponds, where it is also a potential risk for introducing snails and infective cercariae. Hence, it is likely to facilitate the introduction of O. viverrini into aquaculture farms.
A number of significant diseases in the region are caused by the consumption of raw, marinated, or fermented fish products (Grundy-Warr et al., 2012). These include opisthorchiasis and its associated cholangiocarcinoma (Andrews et al., 2008; Sripa et al., 2011), as well as intestinal trematode infections (Chai et al., 2005). Data from this study indicate that both native and aquaculture fish are potential sources of FZT; hence, additional control methods such as HACCP (FAO, 2010) are required for aquaculture fishery.
Conclusion
We have shown that O. viverrini infect aquaculture fish in Thailand, which has not previously been reported. The introduction of PCR in conjunction with conventional methods has shown that O. viverrini as well as other FZT metacercariae infect the most commonly used species of aquaculture fish, B. gonionotus (silver barb) and C. mrigala (mrigal). In contrast to native fish, the aquaculture fish of these and other cyprinid fish are distributed from fisheries to other local and international regions, hence having a high potential for the distribution of FZT parasites and associated diseases (i.e., opisthorchiasis). A limitation of the present study is that it was confined to a small area; comprehensive studies over a large scale are required to determine the extent of O. viverrini and other FZT in aquaculture fish. Such information will enable the development and implementation of suitable strategies to combat the transmission of these foodborne diseases.
Footnotes
Acknowledgments
This research was supported by the Thailand Research Fund through the Royal Golden Jubilee Ph.D. Program (grant PHD/0210/2550 to O.P.), and Higher Education Research Promotion and National Research University Project of Thailand, Office of the Higher Education Commission, through health cluster (SHeP-GMS). We acknowledge the support of the Faculty of Medicine, Khon Kaen University Visiting International Professor Program. We would like to thank the Deutsche Forschungsgemeinschaft (grant PE1611/1-3); the National Research Council of Thailand; ASEAN-EU Year of Science, Technology and Innovation, 2012; and the International Excellence Fund of Karlsruhe Institute of Technology for providing funding for cooperative workshops.
Disclosure Statement
No competing financial interests exist.