
Abstract
Thirty-nine Shiga toxin–producing Escherichia coli (STEC) O113 Irish farm, abattoir, and clinical isolates were analyzed in conjunction with eight Australian, New Zealand, and Norwegian strains for H (flagellar) antigens, virulence gene profile (eaeA, hlyA, tir, espA, espB katP, espP, etpD, saa, sab, toxB, iha, lpfA O157/OI-141, lpfAO113, and lpfAO157/OI-154), Shiga toxin gene variants (stx 1c, stx 1d , stx 2, stx 2c, stx 2dact, stx 2e, stx 2f, and stx 2g) and were genotyped using pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). All of the Irish strains were O113:H4, regardless of source, while all non-Irish isolates carried the H21 flagellar antigen. The stx1 gene was present in 30 O113:H4 strains only, whereas the stx2d gene was common to all isolates regardless of source. In contrast, eaeA was absent, while hlyA was found in the Australian, New Zealand, Norwegian, and two of the Irish human clinical isolates. saa was present in the O113:H21 but not in the O113:H4 serotype. To the best of the author's knowledge, this is the first report of clinically significant STEC lacking both the eaeA and saa genes. PFGE analysis was inconclusive; however, MLST grouped the strains into three sequence types (ST): ST10, ST56, and ST223. Based on our findings, it was concluded that the stx2d gene is common in STEC O113, which are generally eaeA negative. Furthermore, STEC O113:H4 is a new, emerging bovine serotype of human clinical significance.
Introduction
STEC O113 has been frequently isolated from dairy and beef cattle (dos Santos et al., 2010; Fernandez et al., 2010). STEC O113:H-, O113:H4, and O113:H21 have also been isolated from a range of foods, including cheese, game, pork and pork products, beef and beef products, raw milk, and raw spreadable sausage (Werber et al., 2008). Indeed, a recent U.S. ground beef study reported O113:H21 as the most common serotype recovered (Bosilevac and Koohmaraie, 2011).
The first reported clinical cases of STEC O113 were serotype O113:H21 in hemolytic uremic syndrome (HUS) patients in Australia (Paton et al., 1999). More recently, members of this serogroup have been associated with individual cases of diarrheal illness in Ireland (Garvey et al., 2008, 2009a,b). The increased detection of the O113 serogroup and its links to clinical cases highlights the importance of this serogroup as an emerging pathogen.
STEC are characterized by the production of a range of Shiga toxins (also known as verocytotoxins), which can be differentiated into two major groups (stx 1 and stx 2). The stx 1 group is quite homogenous, consisting of two variants: stx 1c (Zhang et al., 2002) and stx 1d (Kuczius et al., 2004). In contrast, the stx 2 group is much more heterogeneous with variants: stx 2c, (Hii et al., 1991) stx 2d (Piérard et al., 1998), stx 2dact (Melton-Celsa et al., 1996), stx 2e (Muniesa et al., 2000), stx 2f (Schmidt et al., 2000), and stx2g (Leung et al., 2003). STEC may also harbor a number of other virulence genes, including those required for the formation of attaching and effacing (A/E) lesions (Jerse et al., 1990; Blanco et al., 2006), the genetic determinants of which are located on the locus of enterocyte effacement (LEE) (Bolton, 2011).
LEE genes include eaeA (intimin) and tir (EspE; translocated intimin receptor), espA, espB, espD, espF, espG, and espH (type III secretion system [TTSS]) (Bolton, 2011). Other chromosomally located genes include the long polar fimbriae (lpf)–encoding genes (Torres et al, 2002). lpfAO113 is particularly associated with the LEE-negative serotype O113:H21 and the adherence-mediating iha gene (Tarr et al., 2000; Schmidt et al., 2001). STEC may also carry virulence plasmids such as pO157 which contain virulence-associated genes (McNally et al., 2001), including the EHEC-enterohemolysin gene (hlyA), the extracellular serine protease gene (espP), the catalase peroxidase gene (katP), the type III secretion pathway–associated gene etpD, and the adherence-associated gene toxB (Bolton, 2011). Another virulence plasmid pO113 (Paton and Paton, 2002) encodes the STEC autotransporter (sab), which has been shown to promote adherence to human epithelial cells and mediate biofilm formation (Herold et al., 2009).
Recent studies on non-O157 STEC from farms and abattoirs in Ireland reported the serotype O113:H4 as being ubiquitous in both environments (Monaghan et al., 2011). Furthermore, this serogroup has become more prevalent in human clinical cases over the last 5 years. The objective of this study was to characterize Irish farm, food, and clinical O113 isolates in terms of virulence profile and genotype, and to compare them with a representative selection of food and clinically significant isolates from Norway, Australia, and New Zealand.
Materials and Methods
Isolate collection
Irish isolates from bovine feces (27), hide (5) carcass (1), and beef farm soil (4) (Table 1) were obtained from the Teagasc Food Research Centre (Ashtown) culture collection. The fecal isolates were originally from four different farms and isolated on six different sampling trips; the soil isolates were all from the same farm, but isolated on four different sampling occasions (Monaghan et al., 2011). The hide and carcass isolates were from the same abattoir, isolated on the same day with the exception of one of the hide isolates, which was from a different abattoir (Monaghan et al., 2012). The two Irish clinical isolates were provided by the Public Health Laboratory (PHL HSE DML, Cherry Orchard, Dublin). Both strains (Ire 1 and Ire 2) were isolated from patients with mild gastroenteric illness. Isolates from Australia and New Zealand were kindly provided by Prof. James Paton and Dr. Adrienne Paton (University of Adelaide). The former included clinical strains (HUS [AUS 1]), microangiopathic hemolytic anemia (MAHA) and thrombocytopenia (Aus 3), and an O113:H21 outbreak (asymptomatic carrier [Aus 5]) and a fermented sausage isolate (AUS 2). Both New Zealand strains (NZ 1 and NZ 2) were isolated from HUS patients, while the Norwegian isolates (Nor 1 and Nor 2) were obtained from bovine fecal samples and kindly provided by Prof. Yngvild Wasteson of the Norwegian School of Veterinary Science.
HUS, hemolytic uremic syndrome.
Serotyping
All isolates were serotyped at the Pennsylvania State University (University Park, PA), as previously described (Monaghan et al., 2011, 2012).
Detection and of virulence genes and variants
All isolates were screened for the presence of stx 1, stx 2, eaeA, hlyA, tir, espA, katP, espP, etpD, hlyA, espB, saa, sab, toxB, iha, lpfA O157/OI-141, lpfA O113, and lpfA O157/OI-154, as previously described (Monaghan et al. 2011). The Shiga toxin genes were subtyped using previously published typing protocols and conditions (Table 2). A list of positive control strains and references is provided in Table 3.
HC, hemorrhagic colitis; HUS, hemolytic uremic syndrome; CDC, Centers for Disease Control and Prevention.
MLST analysis
Seven housekeeping genes were amplified and sequenced in accordance with the protocol of the Escherichia coli Multilocus Sequence Typing (MLST) Database (
From
MLST, multilocus sequence typing.
All PCR-generated DNA templates were purified using the Qiagen QIAquick PCR purification kit (model 28106), and double-stranded sequencing was carried out by Eurofins MWG (Ebersberg, Germany) along with each respective primer set. All DNA sequence analysis was carried out using programs from the DNASTAR Lasergene 7 software package (DNASTAR, Madison, WI). Assembly was carried out using SeqMan, trimming using SeqBuilder, and alignment using MegAlign through the clustal W method. Each of the seven gene loci was assigned an allele number by submission of the sequences to the MLST Database at the ERI, University College Cork (
PFGE analysis
Pulse field gel electrophoresis was preformed using the standardised protocol for subtyping of E. coli O157:H7, as established by the Centers for Disease Control and Prevention (CDC, Atlanta, GA) (Ribot et al., 2006). Restriction digestion was carried out on 2-mm slices of plugs by incubating overnight with the restriction enzyme XbaI (New England Biolabs, Ipswich, MA). Gels were immersed in O.5×Tris/boric acid/EDTA buffer (TBE; Sigma-Aldrich, St. Louis, MO) and run with the following parameters: initial switch time 2.16 s, final switch time 54.17 s, run time 18 h, 30-kb low MW, 600-kb high MW, temperature 14°C, ramping factor linear, pump at 11 min−1. PFGE profiles were normalized and subjected to computer-assisted DNA fingerprint analysis using BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium). The genetic diversity and relatedness of the E. coli O113 isolates were compared at an 80% similarity criterion using Dice coefficient and the underweighted pair group method (UPGMA).
Results
Serotypic analysis of the O (lipopolysaccharide) antigens confirmed all the tested isolates as O-type 113. H (flagellar) analysis identified the Irish isolates (clinical, farm and abattoir) as H type 4 and the remaining (Australian, New Zealand, and Norwegian) isolates as H type 21.
The distribution of virulence genes detected in the isolates is shown in Table 5. The toxin gene stx 2d was common to all isolates (100%), while stx 1 was present in the majority of farm, abattoir, and clinical isolates of Irish origin (83%). The stx 1c gene was present only in the two Irish human clinical isolates. The attachment-associated gene iha was present in all isolates, and the enterohemolysin gene hlyA was common to all clinical, food, and Norwegian bovine isolates. The plasmid-encoded espP gene was also detected in these isolates and in addition was detected in one Irish bovine hide isolate (23% of all isolates). The pO113-associated lpfAO113 and saa genes were present in all isolates from Norway, Australia, and New Zealand (17% of all isolates), while the sab gene was only detected in the clinical isolates from Australia and New Zealand (11% of all isolates). All isolates lacked the following genes: eaeA, tir, espA, espB, katP, etpD, toxB, lpfAO157/OI-141, and lpfAO157/OI-154.
Note: eaeA, tir, espA, espB, katP, etpD, toxB, iha, lpfAO157/OI-141, and lpfAO157/OI-154 were not detected in any of the isolates.
Thirty-three distinct PFGE patterns were identified from the 46 isolates (Fig. 1). Four different patterns (A–D) contained two or more isolates with “indistinguishable” patterns in accordance with Tenover's interpretation (Tenover et al., 1995); all of these were within the Irish cohort of isolates. The largest of these (pattern A) contained 11 Irish isolates (Ire 153, 11, 37, 140, 35, 36, 143, 104, 105, 81, and 106) with an indistinguishable pattern. These Irish isolates were composed of seven bovine feces and four soil isolates, from three different farms and isolated on different sampling occasions over the course of 12 months (Monaghan et al., 2011). Ire 9 and Ire 99 were closely related to each other and to pattern A. These isolates originated from different farms and were isolated at different times of the year. Pattern B comprised two “indistinguishable” isolates (Ire A7 and A8 bovine hides) which were also “closely related” (within 2–3 fragment differences) to Irish isolates Ire A10 and A9 (bovine hides). These four isolates were obtained in the same abattoir on the same occasion. Pattern C comprised two “indistinguishable” isolates (Ire 161 and 162) which were from bovine feces and were isolated from the same farm on the same sampling trip. Pattern D comprised two “indistinguishable” isolates (Ire 157 and 159) originating from bovine feces; these isolates also originated from the same farm and were isolated on the same sampling trip. All other 25 PFGE patterns were unrelated and distinct from each other. This included all the human clinical isolates from Ireland, Australia, and New Zealand and the food isolate from Australia and the bovine isolates from Norway.
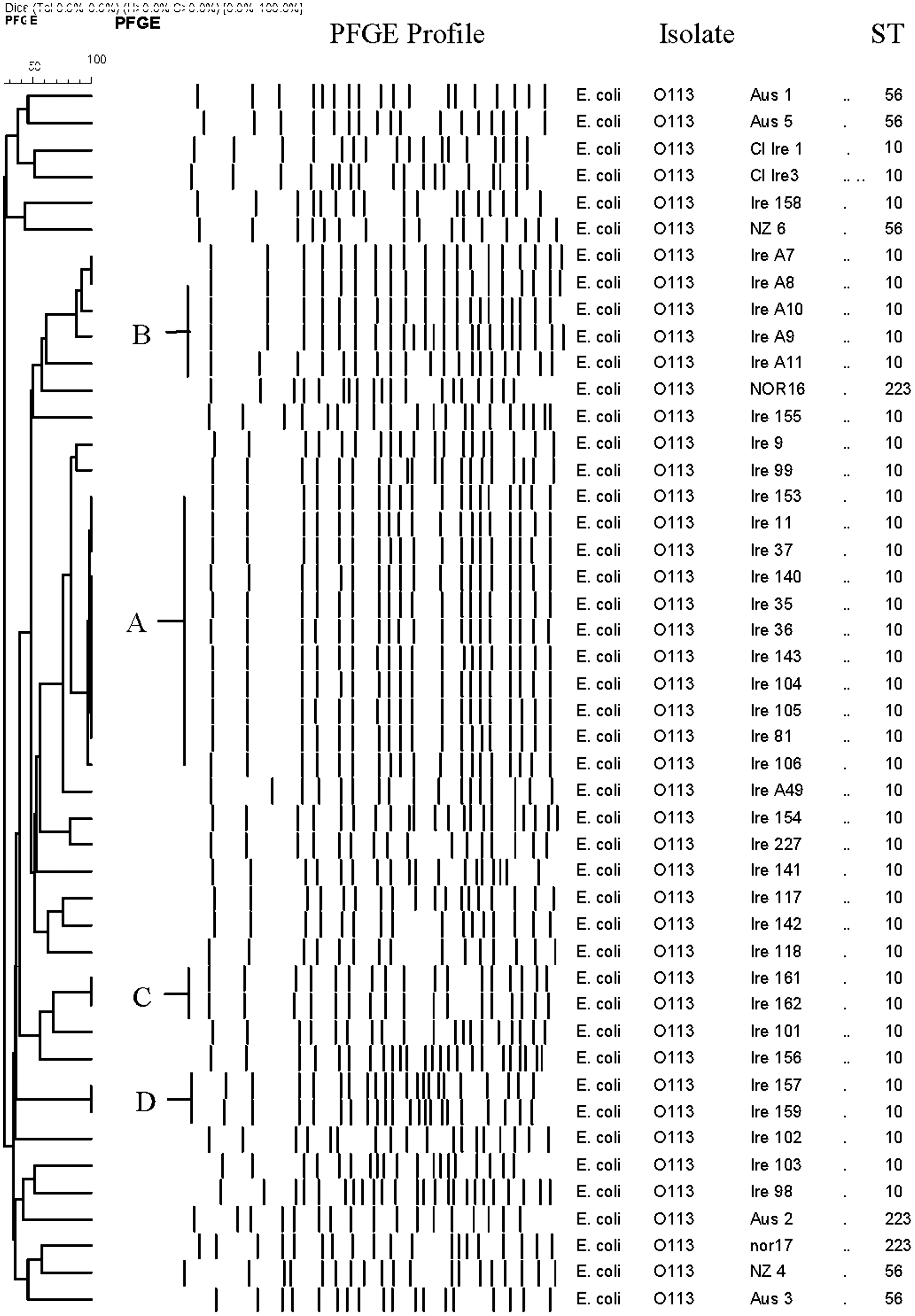
Pulsed-field gel electrophoresis (PFGE) profile of farm, abattoir, food, and clinical O113 isolates.
The 46 isolates provided seven MLST loci and could be grouped into three different STs (Fig. 1). All of the Irish isolates belonged to ST 10. The Norwegian bovine isolates and the Australian food isolate belonged to ST223, and the clinical isolates from Australia and New Zealand all belonged to ST56.
Discussion
All of the Irish STEC O113 isolates were H type 4, regardless of source (farm, abattoir, or human clinical). This serotype has been previously isolated from foods of bovine origin and has been implicated in human illness (Prager et al., 2005; Werber et al., 2008). The O113:H4 predominance in Ireland is in contrast to other countries where H type 21 is more common as a causative agent in human infections (Paton and Paton, 1999; Hogan et al., 2001; Blanco et al., 2003).
Virulence gene analysis of the isolates showed that stx 2d was common to all, regardless of source or geographic origin. The presence of this variant across all of the O113 isolates correlates with the findings of previous work (Miko et al., 2009). A close association has been found between the severity of disease in infected patients and the presence of the stx 2 gene (Beutin et al., 2002). The stx 1 gene was present in the majority of the Irish isolates, and the human clinical strains carried the stx 1c variant. This was not unexpected as the stx 1c variant is frequently detected in O113:H4 (Friedrich et al., 2003) and is commonly associated with intimin-negative isolates.
In addition to Shiga toxin–producing genes, virulence gene profiling data obtained in this study also suggested that a number of other genes (hlyA, espP, and iha) are important in O113-associated illness in humans. These genes were common to all clinical isolates regardless of source or geography. In fact, the iha gene was present in all isolates including those of clinical origin. It has previously been described as an adhesion factor and has been shown to increase adhesion to epithelial cells (Leveile et al., 2006). The putative adhesion gene saa was present in the clinical isolates from Australia and New Zealand, but this gene was absent from the Irish clinical isolates. It has been previously speculated that the presence of the saa gene is a mandatory factor in the facilitation of human illness with STEC O113 (Paton and Paton, 1999). However, our data clearly demonstrates that the saa gene was absent in strains associated with gastrointestinal illness, and to the best of our knowledge this is the first report of a clinically significant STEC strain lacking both saa and eaeA genes.
The hlyA and espP gene products may also play a significant role in pathogenicity. Both genes were exclusive to the clinical isolates with the exception of one abattoir isolate which carried the latter. It is generally accepted that the hlyA gene product has a key role in human infection. Antibodies to this hemolysin have been detected in the sera of convalescent HUS patients (Schmidt et al., 1995), and a cytotoxic effect of the HlyA toxin on human endothelial cells has also been reported (Aldick et al., 2007). The espP gene has similarly been linked to pathogenic STEC (Jarvis and Kaper, 1996; Brunder et al., 1997).
The LpfA genetic variant lpfAO113 was not detected in any of the Irish isolates regardless of source. This gene was however present in the clinical isolates originating from Australia and New Zealand. The absence of this gene in (Irish human) clinical isolates suggests that its products are not essential for associated illness in humans and may be exclusively affiliated with O113:H21 isolates.
MLST analysis assigned the isolates to three STs. All of the Irish isolates tested were assigned to ST10; all of these isolates shared similar virulence profiles and were serotype O113:H4. Five isolates (all clinical isolates from Australia and New Zealand) were assigned to ST56; all isolates shared identical virulence profiles and were O113:H21. The remaining isolates (three) were assigned to ST223 (Norwegian and Australian); all of these were O113:H21 isolates with identical virulence profiles. Of the STs identified, ST10 and ST223 have been previously isolated from humans (E. coli MLST Database,
PFGE clustered the Irish isolates of farm and abattoir origin together with exceptions. The Australian clinical isolates Aus 3 and Australian food isolate Aus 2 clustered away from the rest of the clinical isolates. The remaining Australian, New Zealand and Irish clinical isolates and Norwegian bovine isolates clustered together. These groupings were in contrast to the MLST findings and illustrate that, although they may be viewed as complementary techniques, discrepancies between molecular typing methods remain (Melles et al., 2007).
The PFGE “indistinguishable” patterns were confined to Irish nonhuman isolates only. Evidence of clonality was found in Irish bovine feces and soil. This is as expected, considering the cattle grazing routines and associated fecal contamination of the Irish pastures. The absence of a PFGE link between the two Irish human isolates suggests that these infections may have been sporadic and may not be associated with the Irish bovine or environmental isolates.
Conclusion
This study demonstrated the transfer of geographically similar STEC O113 from soil, bovine feces, and hides to carcasses. Virulence profiling suggests the farm and abattoir isolates were potentially clinically significant. When comparisons were carried out between the Irish clinical and non-clinical isolates, virulence gene profiling was found to be very similar with all isolates carrying stx 1, stx 2d, and iha genes. The Irish isolates had the same ST using MLST analysis; however, further discriminatory PFGE typing did not show clonality between the Irish human isolates and the bovine or environmental isolates. The Irish clinical isolates also lacked the saa gene, and this demonstrated, for the first time, that isolates which lack the eae and saa genes may also be clinically significant, suggesting another mechanism of attachment. The Australian, New Zealand, and Norwegian isolates showed fewer similarities to both the clinical and environmental Irish isolates, with the most notable difference being their H type and “different” PFGE patterns.
The findings of this work emphasize the need for continued monitoring of non-O157 serogroups, especially within the food chain, where the focus of monitoring is heavily biased toward O157. Furthermore, STEC O113 is emerging and may be a significant threat to public health in the future. Further research on the epidemiology and pathogenicity of this serotype is therefore urgently required.
Footnotes
Acknowledgments
This project was funded by the Department of Agriculture, Fisheries and the Marine (DAFM) under the Food and Institutional Research Measure (FIRM), Ireland.
Disclosure Statement
No competing financial interests exist.