
Abstract
In this study, the inhibitory efficacy of Rosmarinus officinalis essential L. (ROEO) and 1,8-cineole (CIN) in inhibiting the growth and survival of Staphylococcus aureus ATCC 6538 and the induction of direct and bacterial cross protection (lactic acid pH 5.2; NaCl 100 g/L; high temperature 45°C) were evaluated following exposure to sublethal and increasing amounts of these treatments in meat broth. All of the concentrations of the ROEO and CIN examined in this study (minimum inhibitory concentration [MIC], 1/2 MIC, and 1/4 MIC) inhibited the viability of S. aureus throughout the 120 min of exposure. The overnight exposure of S. aureus to sublethal amounts of both ROEO or CIN in meat broth did not result in direct or cross protection. Cells progressively subcultured (24-h cycles) in meat broth with increasing amounts of ROEO or CIN showed no increased direct tolerance. These results reveal the antimicrobial efficacy of ROEO and CIN for use in food conservation systems as anti–S. aureus compounds given their efficacy at inhibiting bacterial growth, in addition to their lack of induction for the development of homologous and heterologous resistance.
Introduction
Some researchers have found resistance in isolates of S. aureus from foods to some antimicrobials used in food conservation and sanitization (Carson et al., 2002; Severin et al., 2004). Exposure of microorganisms to sublethal concentrations of inhibitory compounds may lead to adaptation to the same or cross tolerance to a range of unrelated stressing agents (Hill et al., 2002). The assessment of the response to the sublethal injuries in food-related microorganisms could be helpful for reducing the uncertainty about the establishment of the true microbiological risk related to the lack of knowledge about the physiological behavior of the microbial cells surviving in sublethal stress (Luz et al., 2012).
Concern over the negative consumer perception of chemical preservatives has increased interest in the use of natural compounds to control the growth and survival of bacteria in foods (Barros et al., 2009). Early studies showed that Rosmarinus officinalis L. essential oil (ROEO) possesses strong and wide-spectrum antimicrobial activity against spoilage and pathogenic food-related bacteria (Santoyo et al., 2005; Prabuseenivasan et al., 2006). The antimicrobial property of ROEO has been mainly related to the monoterpene 1,8-cineole (CIN), which has been often found as its majority compound (Naveena et al., 2006).
This study assessed the efficacy of ROEO and its main component CIN in the growth inhibition and survival of S. aureus ATCC 6538, and evaluated the development of direct and/or cross protection when the strain was exposed to sublethal concentrations of these substances in food-based media.
Materials and Methods
Essential oil and 1,8-cineole
ROEO (batch ROSTUN04; density at 20°C, 0.94; refractive index at 20°C, 1.51) obtained by steam distillation was purchased from Aromalândia Ind. Com. Ltda. (Minas Gerais, Brazil), and its quality parameters were described in an accompanying technical report. CIN was purchased from Sigma Aldrich (Sigma, Lyon, France). A previous study (Azerêdo et al., 2011) found CIN (32.2 g/100 g) as the majority compound of the ROEO examined here. Solutions of the antimicrobials were prepared in nutrient broth (Himedia, Mumbai, India) in a range of 160 to 0.075 μL/mL using bacteriological agar (0.015 g/L) as a stabilizing agent (Bennis et al., 2004).
Test microorganism
S. aureus ATCC 6538 was obtained from the Collection of Reference Microorganisms, National Institute of Control Quality in Health (FIOCRUZ, Rio de Janeiro, Brazil). A stock culture was kept on nutrient agar (Himedia, Mumbai, India) under refrigeration (4±1°C). Unless stated otherwise, the inocula [approximately 10 log of colony forming units per milliliter (log CFU/mL)] used in the assays were obtained from stationary phase suspensions of the strain and prepared according to a previously described procedure (Carson et al., 2002). The obtained suspension was serially diluted (10−1 to 10−3) in 0.1 M phosphate-buffered solution [PBS (NaCl 9 g/L; pH 7.4)] to provide a viable cell count of approximately 7 log CFU/mL.
Preparation of meat broth
Time-kill and assays to determine the development of direct and cross protection were carried out using cultures grown in a meat-based broth as the substrate for bacterial cultivation. To generate this broth, bovine meat steaks were trimmed of all external fat and cut into uniform-sized pieces (3×3×3 cm). Meat pieces were then boiled in distilled water for 20 min at 90°C. Approximately 500 mL of meat broth was obtained and vacuum filtered using Whatman no. 1. The filtrate was sterilized by autoclaving for 15 min (1.21 atm) and stored at −20°C in aliquots of 50 mL. For use in the functional assays, one aliquot was thawed at a time under refrigeration (7±1°C) (Oliveira et al., 2010).
Determination of the minimum inhibitory concentration
Minimum inhibitory concentration (MIC) values of ROEO and CIN were determined using macrodilution in broth. Double strength nutrient broth (4 mL) was inoculated with 1 mL of the bacterial suspension, mixed with 5 mL of the antimicrobial solution (160 to 0.075 μL/mL) and vortexed for 30 s. The system was statically incubated for 24 h at 35°C. The MIC was defined as the lowest concentration of ROEO or CIN that was required to prevent visible bacterial growth (Nostro et al., 2001). Control flasks without the antimicrobials were tested similarly.
Time-kill assay
The effect of ROEO and CIN (MIC, 1/2 MIC, and 1/4 MIC) on the viability of S. aureus in meat broth over a time span of 120 min was evaluated by a viable cell count procedure. Meat broth (4 mL) was inoculated with 1 mL of the bacterial inoculum, 5 mL of the antimicrobial solution was added to the system, and the culture was gently shaken for 30 s using a vortexer. The system was incubated at 35°C, and at increasing time intervals (0, 15, 30, 45, 60, and 120 min) 1 mL of the suspension was serially diluted (10−1 to 10−5) in PBS, spread on sterile nutrient agar, and allowed to grow for 24 h at 35°C (Nostro et al., 2001). Control flasks without ROEO or CIN were tested similarly. The results were expressed in measurements of log CFU/mL.
Evaluation of the occurrence of bacterial direct protection
Bacterial strains were exposed overnight to sublethal amounts of ROEO or CIN in meat broth according to a previously described procedure (Luz et al., 2012). Bacterial suspensions (1 mL) were inoculated in 20 mL of meat broth, the ROEO or CIN was added (final concentrations of 1/2 MIC or 1/4 MIC), and the cultures were shaken for 30 s using a vortexer (adaptation treatment). Control flasks without the ROEO or CIN were assayed similarly (non-adaptation treatment). The flasks were incubated overnight (18 h) at 35°C. Following the incubation period, an aliquot (2 mL) of each treatment was used to inoculate fresh meat broth (18 mL) to which the ROEO or CIN (final concentration of the MIC was determined previously) was added. The cultures were shaken for 30 s and incubated at 35°C. Viable cells were enumerated over time (0, 30, 60, 180, and 240 min) by serial dilution (10−1 to 10−5) in PBS, plated on nutrient agar, and grown for 48 h at 35°C. The results were expressed in measurements of log CFU/mL. The induction of bacterial direct protection was determined by comparing the viable cell counts over time in the treated and untreated suspensions upon further inoculation into meat broth, to which the same stress agents were added at their MIC values.
Evaluation of the occurrence of bacterial cross protection
Measurements of cross protection induction were performed according to a previously described procedure (Luz et al., 2012). Bacteria were exposed overnight to sublethal amounts of ROEO or CIN in meat broth, followed by exposure to other stress agents (high temperature, low pH, and NaCl). Preliminary experiments were performed to evaluate the salt tolerance, acid tolerance, and thermotolerance of the bacteria. Untreated cultures were used to inoculate meat broth or meat broth containing NaCl (Qeel Ltda., São Paulo, Brazil; 50–150 g/L, at 35°C), lactic acid (VETEC Química Fina Ltda., Rio de Janeiro, Brazil; pH 4.5–6.0, at 35°C), or meat broth that was incubated at increasing temperature (40–55°C) to determine the NaCl concentration, pH value, and temperature that modestly inhibited cell growth.
After establishing these conditions, an aliquot (1 mL) of a newly obtained bacterial suspension was used to inoculate 20 mL of meat broth, to which the ROEO or CIN was added (final concentrations of 1/2 MIC or 1/4 MIC), and the cultures were shaken for 30 s using a vortexer (adaptation treatment). Control flasks without the ROEO or CIN were assayed similarly (non-adaptation treatment). The flasks were incubated overnight (18 h at 35°C). To evaluate the induction of acid tolerance and osmotolerance, respectively, an aliquot (2 mL) of each sample was used to inoculate fresh meat broth (18 mL) that was acidified with lactic acid to a pH of 5.2 or to which NaCl at 100 g/L was added, and the samples were incubated at 35°C. To measure thermotolerance induction, an aliquot (2 mL) of each treatment was used to inoculate fresh meat broth (18 mL) and incubated at 45°C. Viable cells were determined with increasing time (0, 30, 60, 180, and 240 min) by serial dilution (10−1 to 10−5) in PBS followed by plating on nutrient agar and growth for 24–48 h at 35°C. The results were expressed as log CFU/mL. The induction of bacterial cross protection was measured by comparing viable cell counts with increasing time in the treated and untreated samples following inoculation into growth media either containing stressful additives or exposed to environmental stressful conditions.
Evaluation of the occurrence of the induction of bacterial direct tolerance throughout successive 24-h habituation cycles
The capacity of S. aureus to develop tolerance over a more prolonged exposure time was determined according a previously described method (To et al., 2001; Luz et al., 2012). Bacteria were exposed to increasing amounts of the ROEO or CIN (1/16 MIC, 1/8 MIC, 1/4 MIC, 1/2 MIC, MIC, 2×MIC, and 4×MIC) throughout successive 24-h habituation cycles in meat broth. Specifically, bacterial suspensions (1 mL) were used to inoculate 20 mL of meat broth containing ROEO or CIN (final concentration of 1/16 MIC). The cultures were shaken for 30 s using a vortexer and then incubated for 24 h at 35°C. After the incubation period, an aliquot (1 mL) of each sample was serially diluted (10−1 to 10−5) in PBS and plated on sterile nutrient agar for viable cell determination (35°C for 24 h). At the same time, an aliquot (1 mL) of meat broth to which the ROEO or CIN was added at 1/16 MIC (to allow for bacterial growth) was inoculated in a fresh meat broth to which the same compound at the next highest concentration (1/8 MIC) was added, followed by incubation at 35°C and viable cell detection according to the conditions described above. This procedure was repeated with bacteria exposed to increasing concentrations of ROEO or CIN (1/4 MIC − 4×MIC) until a concentration was reached at which no viable cells were detected.
The detection limit of the method used for viable cell counting was 2 log CFU/mL for all of the assays.
Statistical analysis
The assays were performed in triplicate on two separate occasions, and the results were expressed as an average of the assays. Statistical analysis was performed to determine significant differences (p<0.05) by analysis of variance, followed by Tukey tests using the SigmaStat 3.1 computer program.
Results and Discussion
ROEO and CIN displayed a MIC value of 20 and 40 μL/mL, respectively, against S. aureus ATCC 6538. For all of the concentrations tested (MIC, 1/2 MIC, and 1/4 MIC), the ROEO and CIN inhibited (p<0.05) the cell viability of S. aureus when compared to the control assay. Inhibition of bacterial viability occurred as early as 15 min after exposure, and no subsequent increase in viable counts was observed for the remainder of the exposure. ROEO and CIN at 1/2 and 1/4 MIC decreased the viable count of S. aureus to approximately 5 log CFU/mL after 120 min of exposure, revealing that these concentrations were inhibitory to the growth of the tested strain, but not lethal.
The overnight exposure of S. aureus ATCC 6538 to sublethal amounts (1/2 MIC and 1/4 MIC) of ROEO or CIN did not reveal induction of direct protection (Fig. 1A and B), as demonstrated by the viable cell counts over 240 min of exposure. The kill-curves of S. aureus previously challenged with both tested sublethal amounts of the ROEO presented similar viable counts (p>0.05) when exposed to the same agent at the MIC previously determined (Fig. 1A). Cells that have been previously challenged with sublethal concentrations of CIN displayed a decrease in viable counts (p<0.05) when the cells were further cultivated in growth medium to which the same compound (at its MIC) was added when compared to non-adapted cells (Fig. 1B). However, the cells challenged with sublethal concentrations of CIN displayed a kill-curve shape revealing a decreased relative reduction in the viable cells over 240 min of exposure to the same antimicrobial (at its MIC), while the non-adapted cells displayed a linear and sharp reduction in viable count over the time intervals that were examined.
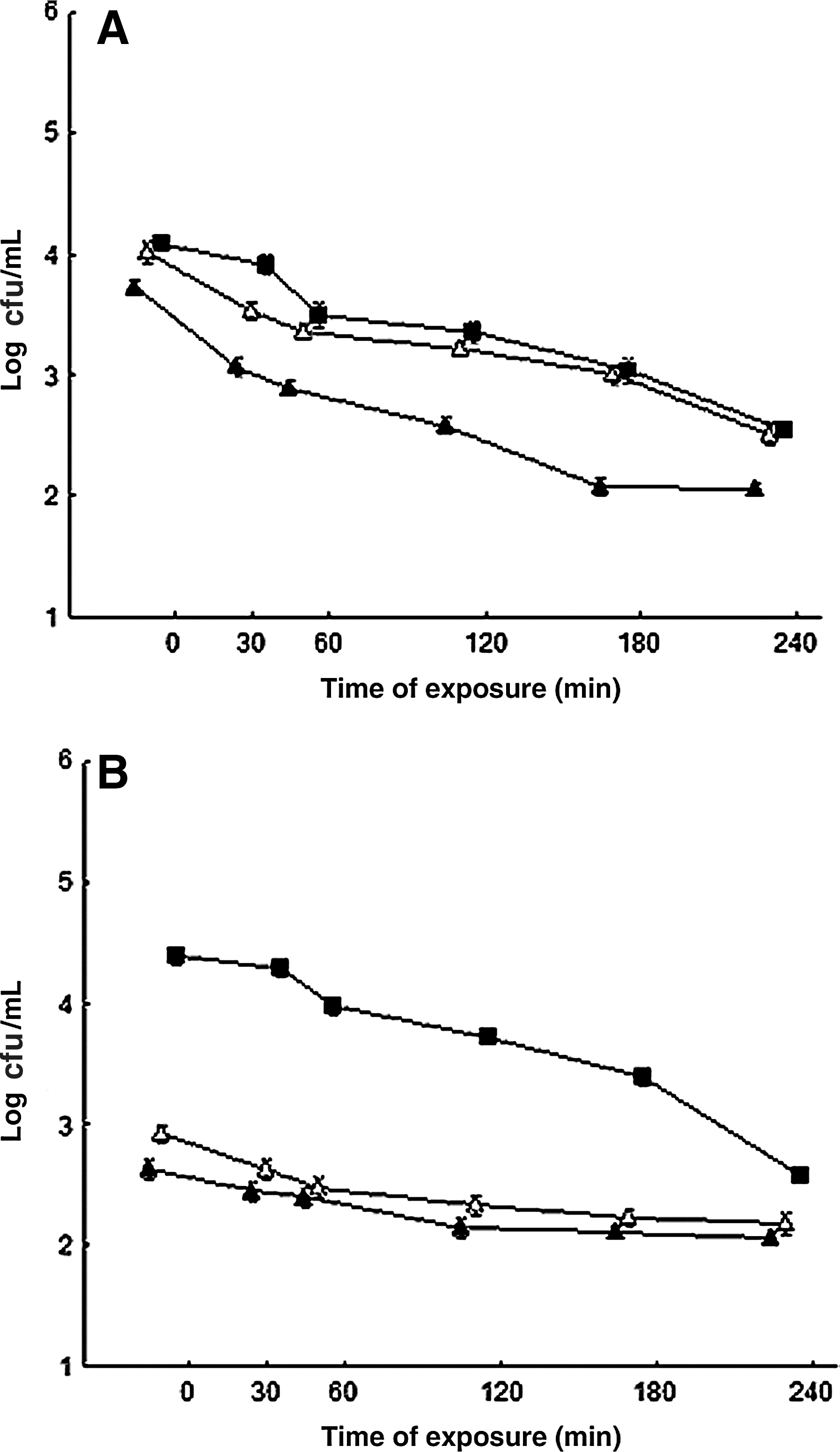
Viable cell counts of Staphylococcus aureus ATCC 6538 in meat broth to which the essential oil and 1,8-cineole (at its minimum inhibitory concentration [MIC]) were added following overnight exposure at 35°C to sublethal concentrations of R. officinalis L. essential oil.
Exposure of S. aureus to sublethal concentration (20 μg/mL) of Epigallocathechin gallate (EGCC), the major phenolic compound of green tea extract, in Muller Hinton broth for 120 min led to adaptation and cross resistance to vancomicyn, ampicillin, and oxacillin with increased MIC values (two- to eightfold increase) (Sing et al., 2007). Still, adaptation to EGCG increased the heat resistance in pre-adapted cells. The authors assumed that exposure of S. aureus to EGCC might cause an increase in the expression of heat shock proteins (HSPs). The HSP DnaK has been shown to play a significant role in the survival of S. aureus under various stress conditions, such as high temperatures, oxidative stress, and in the presence of cell-wall-active antibiotics. HSP is also found in Escherichia coli cells challenged with ethanol, high osmotic pressure, high temperature, and presence of the phenolic carvacrol (Burt et al., 2007).
Consistent with the results of the bacterial direct protection assays S. aureus ATCC 6538 cells that were exposed overnight to sublethal concentrations (1/4 MIC and 1/2 MIC) of ROEO or CIN showed no cross protection to high temperature (45°C), lactic acid (pH 5.2), and salt (NaCl at 100 g/L) (Figs. 2A–C and 3A–C). For the most systems, kill-curves of pre-adapted and non pre-adapted cells revealed similar shapes over the time intervals that were examined. No difference (p>0.05) was found between the counts of viable cells of S. aureus submitted to pre-adaptation with the ROEO or CIN (at 1/2 MIC and 1/4 MIC) upon further exposure to heterologous stressing agents (low pH, NaCl, and high temperature). The cells submitted to pre-adaptation with ROEO or CIN revealed increased sensitivity to lactic acid, high temperature and NaCl when compared to the non-adapted cells, noted for smaller (p>0.05) viable counts over 240 min of exposure.
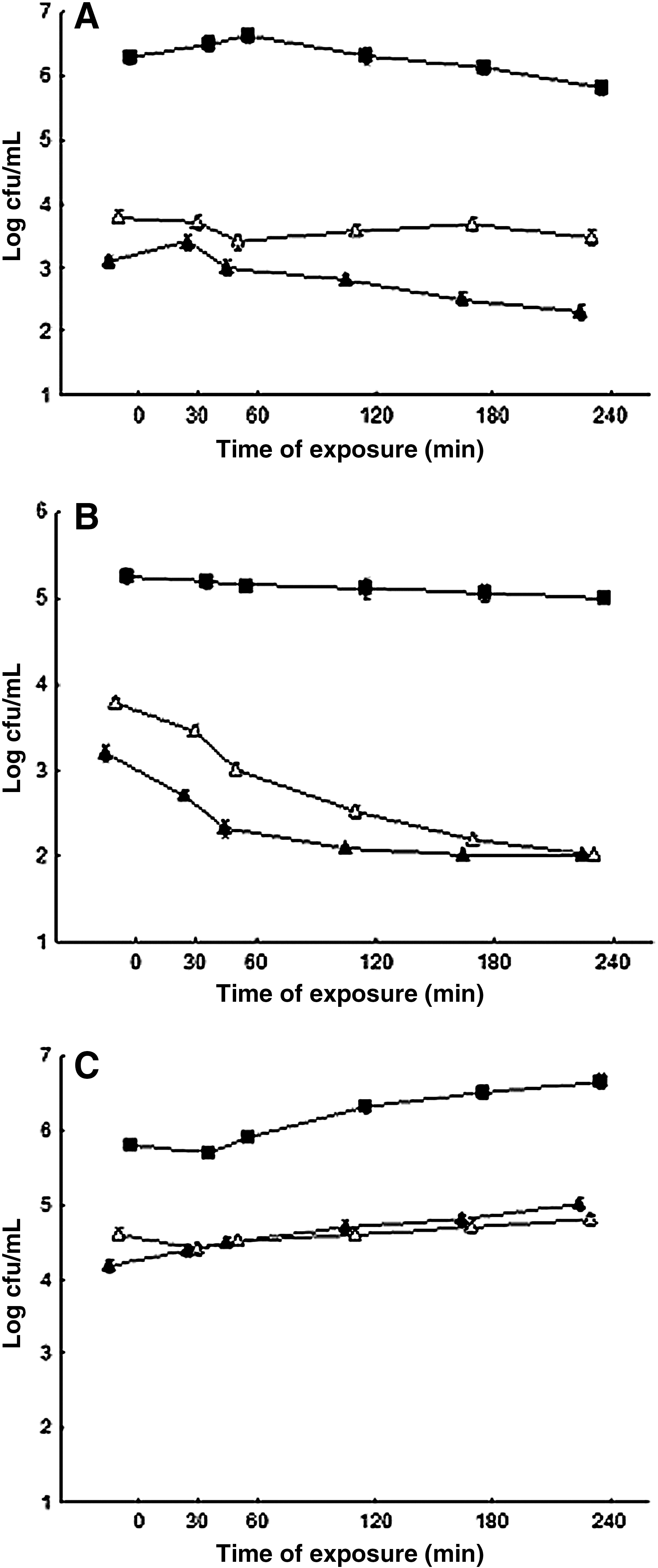
Viable cell counts of Staphylococcus aureus ATCC 6538 grown in meat broth incubated at high temperature 45°C
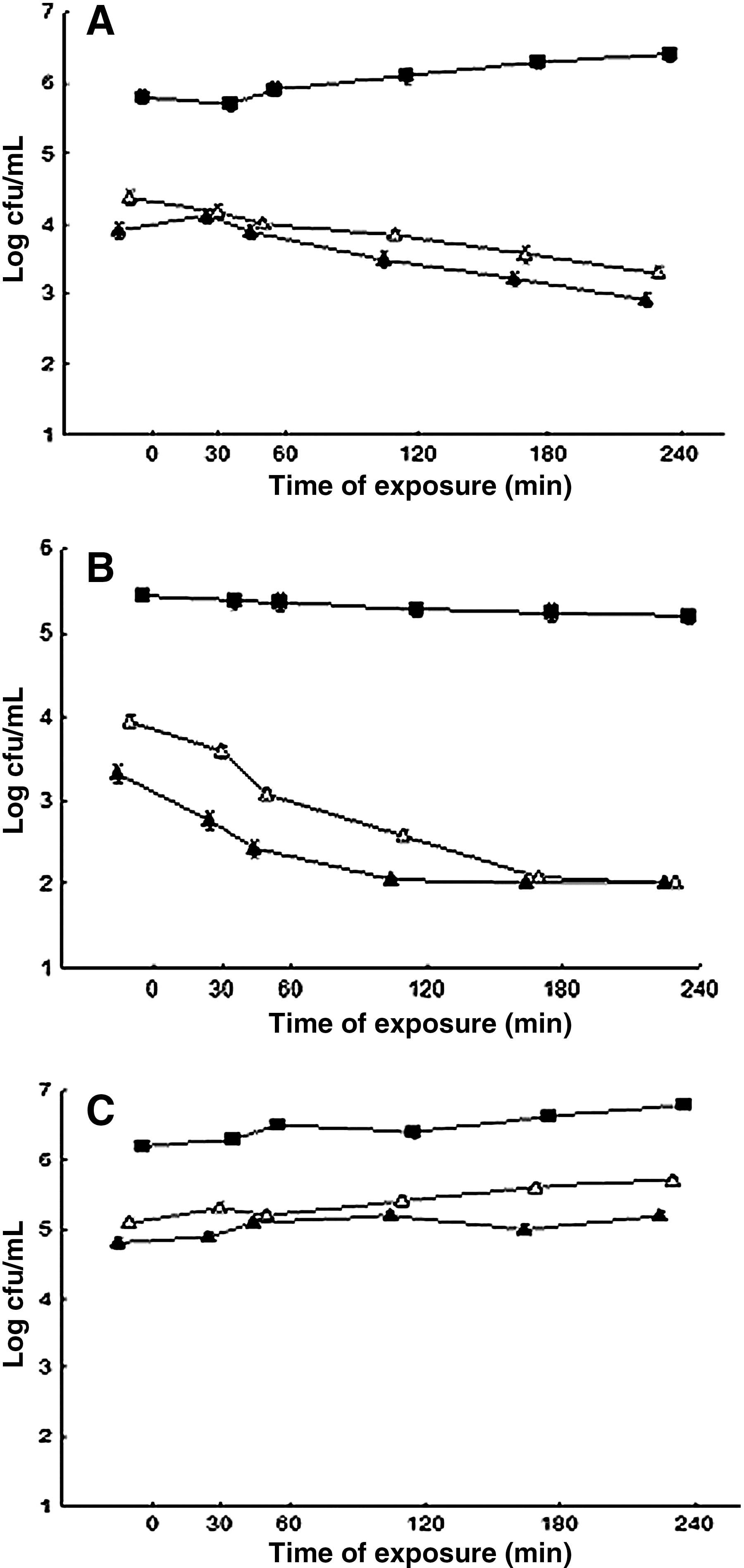
Viable cell counts of Staphylococcus aureus ATCC 6538 grown in meat broth incubated at high temperature 45°C
Cebrián et al. (2010) reported the ability of S. aureus CECT 44459 to develop direct and cross resistance when exposed (5 min to 2 h) to sublethal conditions of acid (hydrochloric acid pH 5.5) and alkaline pH (sodium hydroxide pH 8.0–10.0), hydrogen peroxide (0.01 and 10 mM), and heat (40°C and 48°C) in tryptone soy broth. These researchers noted that the presence of the antibiotics rifampicin and chloramphenicol in the growth media concomitantly to the other stressing agents completely abolished the increase in homologous resistance to pH and hydrogen peroxide. Other studies have also noted that addition of protein synthesis inhibitors, such as rifampicin and chloramphenicol, at different levels in growth media completely prevent the development of resistance to acid pH in bacteria (e.g., Enterococcus faecalis, Lactococcus lactis, Listeria monocytogenes, Salmonella typhimurium), suggesting that protein synthesis is necessary for the development of the resistance responses (Cebrián et al., 2010). Essential oils and their compounds even at levels lower than the MIC have been cited to suppress the synthesis and activity of enzymes in a number of S. aureus strains resulting in protein synthesis block (Nostro et al., 2001; Oliveira et al., 2010), and this could be related to the difficulty of S. aureus ATCC 6538 to develop direct or cross adaptation in the conditions used in this study. Moreover, terpenes found in ROEO (such as CIN, β-pinene, and α-terpinene) have the ability to disturb and penetrate the lipid structure of the bacterial cell wall leading to denaturing of proteins, destruction of cell membrane, cytoplasm leakage, cell lysis, and eventually death. The decrease of pH due to the cell membrane disruption caused by terpenes in bacteria means that control of cell processes such as DNA transcription, protein synthesis, and enzyme activity is lost (Oussalah et al., 2006).
Barros et al. (2009) reported that the cultivation of S. aureus ATCC 6538 in nutrient broth supplemented with sublethal amounts of O. vulgare essential oil (1/2 MIC, 0.3 μl/mL; and 1/4 MIC, 0.15 μL/mL) for 24 h interfered with the metabolic activity of the strain with reduction in salt (NaCl) tolerance, lipase and coagulase activity, and enterotoxin production. The authors stated that the changes in metabolic activity of the bacterium could be related to a sublethal injury of the cell provided by subinhibitory concentrations of the essential oil, which may alter the ability of the cell to osmoregulate adequately, exclude toxic material, and synthesize some physiological determinants, such as proteins and enzymes.
Regarding possible assay limitations for the two sublethal concentrations of ROEO and CIN and the chosen exposition time (18 h) for the pre-adaptation further experiments were performed to assess the induction of tolerance following exposure to 24-h cycles in meat broth to which increasing amounts of the ROEO or CIN (1/16 MIC – 4×MIC) were added. The results of these assays showed that S. aureus ATCC 6538 was able to survive (as demonstrated by viable cell counts) in the broth to which the ROEO or CIN were added in concentrations of up to the respective 1/2 MIC, suggesting that the exposure of the cells to increasing sublethal amounts of the both substances for a more prolonged time (24 h) could not induce the development of adaptive responses towards these compounds. The inhibition of the cells already in the broth added of 1/2 MIC of the tested antimicrobials could be related to the manifestation of injury because when bacterial exponentially growing cells are exposed and continuously kept in a stressing but non-lethal environmental condition, as that provided by the sublethal amounts of the tested antimicrobials, they could lose their viability and capacity to survive over time (Rees et al., 1995). Sublethally injured bacterial cells with the imposed stress caused by levels of antimicrobials lower than the MIC may develop an imbalance between anabolism and catabolism that is sufficient to disrupt the growth and becoming cells unable to form colonies on agar without affecting the metabolic rate, although it can result in self-destruction of the cells through free-radical attack (burst radical production) mainly because of a failure in cell repair mechanisms (Dodd et al., 1997).
The results obtained in this study highlight the antimicrobial efficacy of ROEO and its majority component CIN for use in food conservation systems regarding their efficacy to inhibit the growth and survival of S. aureus, in addition to a lack of development of homologous and heterologous resistance to high temperature, pH, and salt. Further studies are needed to assess the induction of genetic changes, synthesis of stress proteins, and modification of lipid membrane composition in S. aureus after exposure to sublethal amounts of ROEO and CIN, and the occurrence of induction of bacterial tolerance by the tested compounds in S. aureus when cultivated in food matrices.
Footnotes
Acknowledgments
We are grateful to Conselho Nacional de Desenvolvimento Científico e Tecnológico–CNPq (Brazil) for financial support.
Disclosure Statement
No competing financial interests exist.