
Abstract
Non-typhoidal salmonellosis is an important zoonotic disease caused by Salmonella enterica. This work focuses on the identification of Salmonella enterica clonal strains which, presenting a wide distribution potential, express resistance determinants that compromise effectiveness of the antimicrobial therapy. The screening was performed on 506 Salmonella enterica isolates from animals and humans, which were characterized by serovar and phage typing, genome macrorestriction and pulsed-field gel electrophoresis, and detection of phenotypic and genotypic traits for antimicrobial resistance. A Salmonella Enteritidis strain with strong quinolone resistance is spread on three host environments carrying one of the four variants found for the GyrA protein: (1) Asp87Tyr, the major polymorphism found in 39 Salmonella isolates from human origin and six from poultry; (2) Ser83Phe, with four isolates from human origin and one from white stork (Ciconia ciconia); and (3) Asp87Asn or (4) Asp87Gly, with two isolates each from human origins. Several Salmonella Typhimurium strains that presented int1 elements and the classically associated pentaresistance (ACSSuT) phenotype were found distributed between two host environments: domestic animals and humans, domestics and wild animals, or wild fauna plus humans. This study points out the importance of monitoring gut microbiota and its antimicrobial resistance from wildlife, in parallel to livestock animals and humans, especially for animal species that are in close contact with people.
Introduction
S
Salmonella Enteritidis is commonly found in isolates from eggs and broiler meat, with prevalence rates of 0.8% and 5.5%, respectively (Lestari et al., 2009). Human isolates of this serotype are characterized by the highest levels of resistance for nalidixic acid and ciprofloxacin (18.7% and 9.3% in 2010), although frequencies might depend on using Clinical and Laboratory Standards Institute (CLSI) breakpoints or the lower European Committee on Antimicrobial Susceptibility Testing (EUCAST) epidemiological cut-off values (EFSA, 2012). Resistance to ciprofloxacin, a 6-fluoro-quinolone derivative, is clinically relevant since this antimicrobial represents one of the first choices among antimicrobial treatments for severe salmonellosis in elderly or immunosupressed patients (Hopkins et al., 2005). The main mechanisms producing low susceptibility to quinolones are point mutations affecting the quinolone resistance determinant region (QRDR) of the genes encoding gyrase, gyrA and gyrB, or topoisomerase IV, parC and parE, which might have an additive effect, in addition to plasmid-mediated determinants that confer variable resistance and could provide a favorable background for selection of additional traits (Fàbrega et al., 2009).
Salmonella Typhimurium is mostly associated with the consumption of contaminated pork, poultry and bovine meat. These isolates typically share the pentaresistant phenotype ACSSuT (resistance to ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline) often encoded in gene-cassettes linked to mobile elements like SGI1 transposons, integrons, and/or plasmids (Glenn et al., 2011).
Although there have been some human outbreaks due to Salmonella from wild animals (Kapperud et al., 1998; Alley et al., 2002; Pennycott et al., 2006), little is known about the role of wildlife in the spread of antimicrobial resistant bacteria to humans and livestock. The aim of this work is the identification of clonal relationships that allow the tracing and distribution of antimicrobial-resistant strains of Salmonella enterica.
Materials and Methods
Microbiological material
A total of 303 Salmonella isolates were collected in six hospitals of Extremadura (Spain), located in the cities of Cáceres, Coria, Llerena, Mérida, Navalmoral de la Mata, and Plasencia, during the years 2007–2008 (with the exception of 41 strains collected between 2004 and 2006). In parallel, the VISAVET Health Surveillance Centre (Complutense University of Madrid), the Extremadura University, and the Córdoba University provided 203 Salmonella from up to 13 animal species, including wildlife (white stork, black vulture, hawk, partridge, and quail), livestock (white swine, iberian swine, sheep, cow, and horse), and pet animals (python, varano, and canary) that were isolated between 1997 and 2008 in Extremadura and surrounding regions of the southwest of Spain.
Bacterial typing
All the bacteria from animal origin were serotyped, in their laboratories of origin, by slide agglutination with commercial antisera according to the Kauffmann-White scheme. Phage typing was performed according to previously published protocols (Anderson et al., 1977; Ward et al., 1987) with phages and interpreting criteria provided by the Health Protection Agency (HPA, formerly Public Health Laboratory Service, London, UK). Human isolates were serotyped by the same methods and criteria as animal isolates by the Salmonella-Shigella National Reference Laboratory (NRL, National Health Institute Carlos III, Majadahonda, Madrid).
The clonal relationships among the 44 serotypes identified were analyzed by XbaI digestion of total DNA and pulsed-field gel electrophoresis (PFGE) according to PulseNet protocols. A Dice coefficient of 1% was used to construct the relationships dendograms by Bionumerics 5.1 (Bio-Rad).
Antimicrobial susceptibility testing
The minimal inhibitory concentrations (MICs) were determined by Muller-Hinton broth microdilution (ISO 20776-1:2006) for ampicillin, chloramphenicol, ciprofloxacin, nalidixic acid, streptomycin, sulfamethoxazole, and tetracycline, according to the EFSA recommendations (Bronzwaer et al., 2008). The epidemiological cut-off values considered were those recommended by EUCAST (MIC distribution website,
Identification of antimicrobial resistance determinants
Mutations in the quinolone resistance determinant region (QRDR) of gyrA, gyrB, parC, and parE were analyzed by polymerase chain reaction (PCR) (Eaves et al., 2009) and sequencing of the amplified DNA fragments (StabVida, Lisbon, Portugal). Sequences corresponding to plasmid-mediated quinolone resistance determinants qnrA, qnrB, qnrC, qnrD, qnrS, aac(6')-lb-cr, or qepA genes were screened by PCR according to previously reported methods (Catoir et al., 2007; Cavaco et al., 2009; Kim et al., 2009; Wang et al., 2009). Int1 elements and associated gene-cassettes were detected by PCR following described protocols (Leverstein-van Hall et al., 2002).
Results
Clonal analysis of bacterial isolates
The dispersion of Salmonella enterica strains among the three different kind of hosts analyzed in this work—wildlife or domestic animals (livestock and companion animals) plus humans—was evaluated by the clonal analysis of bacteria isolated from every niche (Table 1). Fourteen pulse types belonging to eight different serotypes were shared among isolates found in domestic animals and humans, but not in wild animals. In contrast, five pulse types were detected in isolates from wildlife species; two were found only in one additional host (human or white swine), whereas the remaining three were detected in the three host environments (Table 1).
Pulse types.
Wildlife species are as follows: white stork, Ciconia ciconia; wild boar, Sus scrofa; Partridge, Alectoris rufa.
Domestic species are as follows: canary, Serinus canaria domesticus; cattle, Bos taurus; horse, Equus ferus caballus; poultry, Gallus gallus domesticus; Python, Phyton sp.; white and iberian swine, breeds of Sus scrofa domesticus.
Numbers of isolates > 1 are indicated between parentheses.
Salmonella Typhimurium (97 isolates) and Salmonella Enteritidis (85 isolates) were the most prevalent serotypes among pulse types detected in more than one host environment, with 224 isolates in total. Salmonella Typhimurium (nine pulse types) was more diverse than Salmonella Enteritidis (three pulse types), whereas a single pulse type, EN02, covered the major fraction of isolates from humans (69 isolates) or animals (nine isolates) among the three hosts analyzed in this work (Table 1).
Antimicrobial resistance of widely distributed serotypes
An antimicrobial susceptibility test revealed that most EN02 isolates (56 strains) presented resistance to quinolones, although a reduced group of four isolates was only resistant to nalidixic acid, a phenotype strongly linked to mutations in the gyrA QRDR (Table 2). No other resistance determinant was found, including mutations in the QRDR of gyrB or parE, or plasmid-linked genes like qnrA, qnrB, qnrC, qnrD, qnrS, aac(6')-lb-cr, or qepA, since a mutation found in the QRDR of parC (Thr57Ser) was also found in susceptible strains and thus was considered not associated to resistance phenotypes (M.J. Campos and G. Palomo, personal communication). It is noteworthy that a single phage type (PT), PT1, corresponds to the major fraction of quinolone-resistant EN02 isolates (26/56), carrying Asp87Tyr or Ser83Phe GyrA alleles. The other pulse types of Salmonella Enteritidis that were found among different host environments, EN01 and EN03, were detected with low prevalence and did not share remarkable antibiotic resistance.
Pulse type.
Quinolone resistance; NAL, nalidixic acid; CIP, ciprofloxacin.
Phage type: NT, non-typable; RDCN, react but does not conform.
Numbers of isolates > 1 are indicated between parentheses.
Most but not all Salmonella Typhimurium isolates corresponding to more prevalent pulse types carried int1 elements in association with the common pentaresistance phenotype ACSSuT (Table 3), although resistance to chloramphenicol, streptomycin, and tetracycline were not always observed. Several PS/PT/int1 signatures were detected among different host environments: TY09/104b/int1 (15 isolates from white swine and humans presenting the associated gene-cassette of class A, carrying gene oxa1), TY13/U302/int1 (five isolates, from white swine, human, and horse), TY16/104b/int1 (four isolates from white swine and humans, three presenting the gene-cassette A), TY43/104/int1 (three isolates from canary and humans presenting the gene-cassette B, carrying genes aadA2 and pse1), TY13/203/int1 (two isolates, human and iberian swine), and TY43/U302/int1 (two isolates, partridge and human). Two PS/PT signatures not linked to int1 were also detected among different host environments: TY65/193 (nine isolates from iberian swine and humans) and TY19/195 (four isolates, white stork and humans).
Pulse type.
Pentaresistance profile: A, ampicillin; C, chloramphenicol; S, streptomycin; Su, sulphonamyde; T, tetracycline.
Detection of int1 sequences by polymerase chain reaction.
Phage type: NT, non-typable; PNR, pattern not recognized.
Numbers of isolates and gene cassettes are indicated between parentheses: A, 2.0 kb DNA fragment corresponding to gene oxa1; B, 1.0 plus 1.2 kb DNA fragments, genes aadA2 and pse1; C, 0.8 plus 1.0 kb DNA fragments, genes aadA1 and dhfrVII; Ø, no gene-cassette amplification of int1-positive isolates.
Discussion
The high prevalence of antimicrobial-resistant Salmonella has been a matter of serious concern for public health over the past decade (Lestari et al., 2009; EFSA, 2012). Moreover, although antimicrobials like quinolones to which resistance has been analyzed have had a limited use in animals, resistant traits persist, especially among Salmonella Enteritidis. In addition, this work provides evidence about a possible role of wildlife in the distribution of Salmonella isolates that, having different antimicrobial resistant determinants, share resistance to quinolones (Salmonella Enteritidis) or to the compounds covered by the pentaresistance profile (Salmonella Typhimurium).
Salmonella Enteritidis has been detected in hens, mice, flies, and fecal samples from wild birds of Spanish henhouses, suggesting a role of wildlife as a reservoir for poultry of this serotype (Hernándiz, 2007). However, although quinolone-resistant Salmonella presenting GyrA alleles Ser83Tyr and Asp87Asn have been found in isolates from avian wildlife (Reche et al., 2002), they were not detected in Salmonella Enteritidis, which is the major zoonotic serovar detected in poultry (Lestari et al., 2009). In contrast, closely linked EN02/PT1 quinolone-resistant isolates have been found in this work carrying one of two different gyrA (QRDR) genotypes, Asp87Tyr or Ser83Phe, which presumably arose independently after separation from a common hypothetical ancestor. Both clonally related isolates have been detected on two different host environments, domestic animals and humans (Asp87Tyr genotype), or wild fauna and humans (Ser83Phe), which are partially overlapped by clinical isolates from salmonellosis (Fig. 1). The EN02 strain detected in white stork could have been transferred from human sewage, since the same PS/PT/gyrA genotype was found in human strains (Table 2), although it was not detected in poultry. Nevertheless, this work allows tracing the distribution of a Salmonella Enteritidis strain with a high risk for selection of quinolone-resistance. The EN02/PT1 genetic background could predispose for mutation selection by decreasing to some extent the resistance threshold, like it has been proposed that occurs for bacteria expressing some plasmid-encoded determinants (Fabrega et al., 2009).
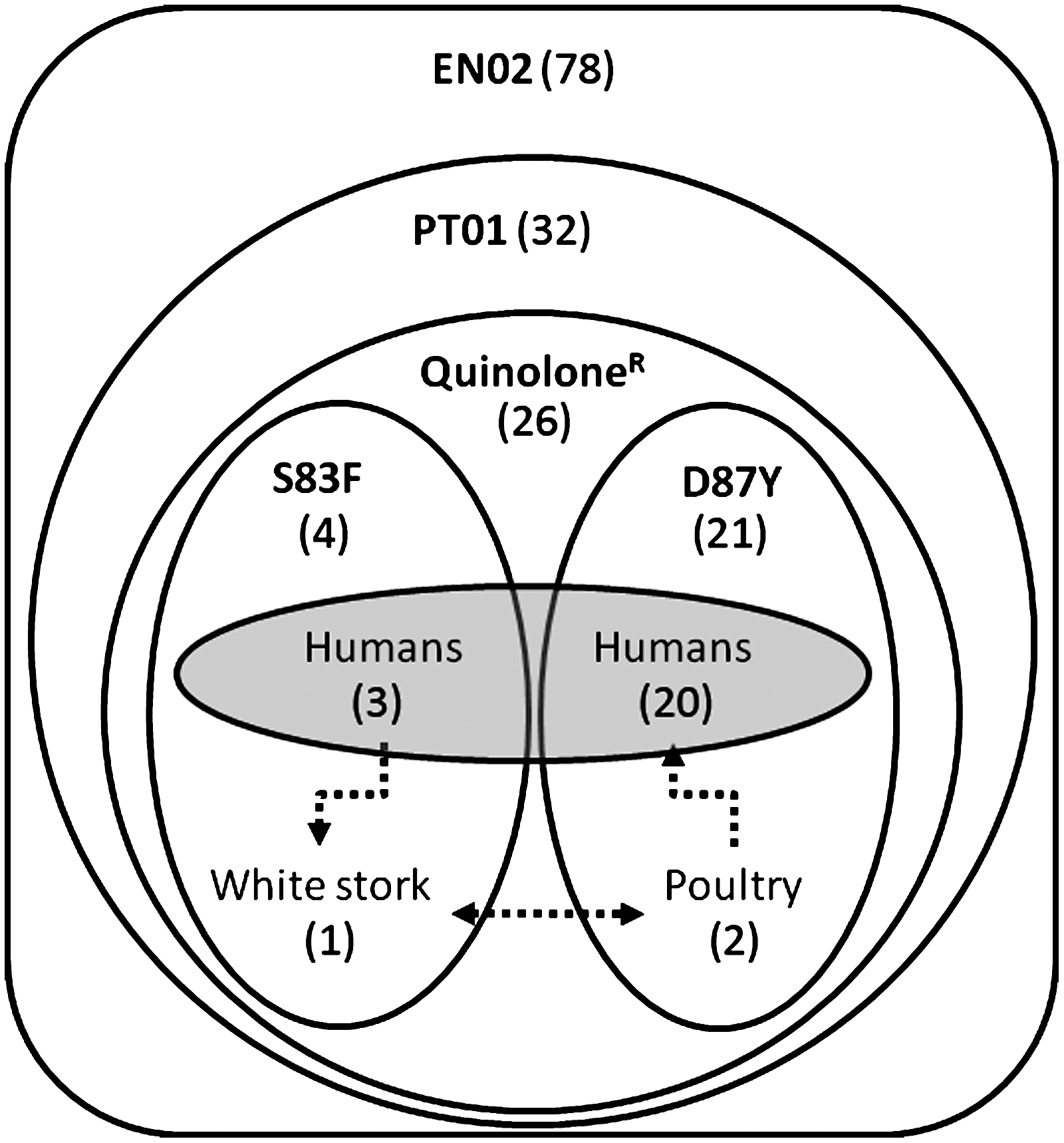
Distribution of Salmonella EN02 isolates among the three host environments. Set of isolates are clustered according to genotypic/phenotypic traits. Numbers of isolates for every category are indicated between parentheses. QuinoloneR, quinolone-resistant strains.
Different studies have described the spread of Salmonella among wild animals from different countries, with Salmonella Typhimurium and Salmonella Enteritidis as the most frequent serotypes (Oloya et al., 2009; Dolejska et al., 2009; EFSA, 2012; Ramos et al., 2010; Taylor and Philbey, 2010). A cross-sectional study performed on Escherichia coli isolations from human sewage, wildlife, domestic animals, farm environments, and surface water indicated that wild animals, in spite of not being naturally in contact with antimicrobials, can serve as reservoirs, vectors, and bioindicators of antimicrobial resistance after been infected with environmental resistant bacteria with origin in humans and livestock (Sayah et al., 2005). To our knowledge, this work is the first that strongly suggests the spread of a quinolone-resistant strain of Salmonella Enteritidis among humans, domestic animals, and wild animals (white stork; Table 2). The clonal diversity displayed by PFGE among the isolates analyzed in this work (M.G. Campos and P. Gonzalo, personal communication) is lower for Salmonella Enteritidis (129 isolates clustered in 24 pulse types) than for Salmonella Typhimurium (189 isolates and 77 pulse types), similar to previous studies (Valdezate et al., 2007), which could contribute to explain, at least in part, the high prevalence of EN02 among other pulse types detected in this work. In addition, PS/PT signatures linked to the ACSSuT pentaresistance profile might target Salmonella Typhimurium strains distributed between two host environments, also including two cases of white stork (Table 3). Isolates lacking int1 might present elements carrying multiple resistance determinants, like IncA/C plasmids (Glenn et al., 2011).
White storks share habitats with people when nesting, feeding, and wintering, from North to South. Their population has increased greatly, mainly due to their free access to dumping sites and to their feeding habits, which include wading in sewage. Although human and stork relationships are clear, its role as a vector of Salmonella to livestock is uncertain, since biosecurity measures keep large wild birds out of intensive swine and poultry farms. However, potential dissemination to developing countries through migration routes could have a strong impact in environments where the control of zoonotic agents is not so effective. The transmission of antimicrobial resistant Salmonella between domestic animals livestock (like poultry and swine), wild fauna, and humans needs further study.
In conclusion, knowledge of Salmonella epidemiology involving humans, livestock, and wildlife is critical to understanding the spread of antimicrobial resistance. Future studies are required to determine if both migrating and waste-feeding would make white stork surveillance worthwhile as a bioindicator of reservoirs of antimicrobial resistant bacteria in wild animals.
Footnotes
Acknowledgments
We wish to thank laboratories of Hospitals "S. Pedro de Alcántara" (Cáceres), "Ciudad de Coria" (Coria, Cáceres), "Campo Arañuelo" (Navalmoral de la Mata, Cáceres), "Virgen del Puerto" (Plasencia, Cáceres), "Mérida" (Mérida, Badajoz), and "Llerena" (Llerena, Badajoz) for providing the Salmonella isolates from human infections; and the Salmonella-Shigella National Reference Laboratory (NRL, National Health Institute Carlos III (Majadahonda, Madrid) for serotyping and phage typing of human isolates. G.P. was supported by a predoctoral fellowship from the Fernando Valhondo Calaff foundation. This work was supported by the Ministry of Education and Science of Spain (project AGL2008-04147/GAN), the Department of Employment, Enterprise, and Innovation of the regional government of Extremadura, Spain (project PRI08B001 and group CTS001) and the University of Extremadura (Group MIVET).
Disclosure Statement
No competing financial interests exist.