
Abstract
Epidemiological data suggests that certain groups such as the elderly are more susceptible to listeriosis than the general population. Repeated exposure to low levels of Listeria monocytogenes may increase the probability of infection. Increased susceptibility to infection in the elderly has been attributed in part to reduced activity of T cells. We investigated the impact of consecutive daily intragastric challenge with L. monocytogenes on the development of listeriosis within an aged guinea pig population. Approximately 63% of animals became infected following oral L. monocytogenes challenge with 104 CFU daily for 3 days, based on recovery of the pathogen from the liver or spleen. At day 4 postchallenge, 100% of animals were infected based on recovery of the bacterium in spleen and liver, decreasing to 14% by day 6 and then steadily increasing to 83% by day 13. During the 13-day postchallenge period, in the blood, numbers of total CD3+ T cells decreased significantly; CD8+ T-cell population underwent two shifts; CD4+ T-cell population decreased and then increased. The results suggest that listerial infection can occur following repeated daily exposure to low levels of L. monocytogenes and that during infection, CD3+ T-cell immune response may be depressed, potentially increasing susceptibility to other diseases.
Introduction
L
Invasive listeriosis predominates in immunocompromised individuals, a group that contributes to the high mortality rate (Wing and Gregory, 2002; Rocourt and Cossart, 2007). Recent epidemiological investigations suggest that ingestion of L. monocytogenes can lead to febrile gastroenteritis without progression to invasive listeriosis. (Aureli et al., 2000; Sim et al., 2002; Danielsson-Tham et al., 2004). The onset of a listerial infection can be sudden, from 9 to 16 hours in healthy individuals manifesting as gastroenteritis, or gradual, from 11 to 70 days, which is typical for invasive listeriosis (Schlech, 1997; Donnelly, 2001; Ooi and Lorber, 2005).
The level of L. monocytogenes reported to cause listerial infection in healthy adults has varied from 105 CFU/g in cold-smoked rainbow trout, 106 CFU/g in salted mushrooms, to 107 CFU/g in smoked mussels (Aureli et al., 2000; Rocourt and Cossart, 2007). For individuals at high risk, the dangerous levels of L. monocytogenes in food ranged from <102 to 104 in rainbow trout to 104 CFU/g in ice cream (Miettinen et al., 1999; Dalton et al., 1997). Actual patient exposure information, single or multiple, associated with a given outbreak is often not available. An outbreak of listeriosis in Finland among immunocompromised patients was associated with prolonged daily consumption of contaminated butter during the hospital stay (Maijala et al., 2001). The level of L. monocytogenes consumed based on levels detected in wholesale butter samples may have been 2.2×104 to 3.1×105 CFU/day. The authors concluded that low levels of L. monocytogenes associated with foodstuff typically used for a longer time can be hazardous for the health of at-risk populations. The severity of L. monocytogenes infections and susceptibility of the elderly is underscored by the recent outbreak of listeriosis in the United States linked to the consumption of contaminated cantaloupe, resulting in 146 persons becoming infected, 33 deaths, and one miscarriage. The median age of confirmed cases was 78, and 88% (129 persons) of cases were ≥60 years old (
Guinea pigs are often used as the animal model to study listerial infection, since listerial challenge by the oral route in mice and rats is not reproducible, and does not become systemic. However, it can be induced in guinea pigs, because L. monocytogenes can cross the intestinal barrier and disseminate throughout the body. The internalization of L. monocytogenes in humans is specific, relying on the interaction between human E-cadherin and internal A (InlA) of L. monocytogenes (Mengaud et al., 1996). The ligand–receptor interaction is species specific; humans and guinea pigs have the same crucial amino acid, proline, at the 16th position (Lecuit et al., 1999). L. monocytogenes is able to cross the intestinal barrier by internalin A-E-cadherin interaction in the guinea pig.
In this article, infection rate and severity of infection based on microbiological analysis of tissue samples following repeated oral exposure to L. monocytogenes (104 CFU) is discussed. Aged guinea pigs (retired breeders) were used to represent an at-risk group for listerial infection (Wu et al., 2012). The immune system response to L. monocytogenes was gauged by following T-cell populations prior to and after challenge with the pathogen.
Materials and Methods
Bacteria
A cocktail of three L. monocytogenes serotype 4b strains (J1-110 [food epidemic], N1-225 [human epidemic], MMS97-1 [raw beef]) were used. Each isolate was cultured individually in 10-mL brain heart infusion (BHI) broth (Difco, Franklin Lakes, NJ) and incubated at 37°C with agitation (200 rpm) for 18 h. Cells were harvested by centrifugation (5500×g, 4°C, 10 min) and resuspended in 10 mL phosphate buffered saline (PBS; Sigma, St. Louis, MO) adjusted to pH 7.2. The cocktail was made by mixing equal volumes of each strain and then serially diluting in PBS to achieve 104 CFU/mL. Cell numbers were confirmed by plating on BHI agar. Stock cultures were maintained in BHI containing 20% glycerol and stored at −80°C.
Animals
Outbred Dunkin-Hartly female retired breeder guinea pigs weighing approximately 1000 g were purchased from Charles River Lab Inc. (Wilmington, MA). All animals were housed individually in separated cages under standard conditions (50% humidity and 12-h dark–12-h light cycle) at the Rutgers animal facility. General Purina guinea pig chow (diet #5025) and fresh water were available ad lib. The feed was stored at room temperature and under dry conditions. Upon arrival, all animals were weighed, and a total of 16 animals were used in each experiment. The experiment was conducted twice. All animal experiments were conducted in accordance with federal guidelines and were approved by the Rutgers University animal care and facilities committee.
Blood and plasma samples collection
Blood samples were collected once a week during the prechallenge period (wk 0–wk 5) and on 4, 6, 8, 11, and 13 days postchallenge. During the prechallenge period at 7-d intervals, five animals randomly selected were anesthetized with ketamine at 80 mg/kg of body weight and xylazine at 20 mg/ kg of body weight. A 1-mL blood sample was collected from the peripheral jugular vein of each animal. Blood samples were collected into syringes containing 5% K2EDTA, and used within 4 h for indirect immunostaining. Blood samples were collected from three animals on each postchallenge day.
Indirect immunostaining
Blood samples were collected for immunostaining in order to determine the levels of target T cells (CD3+, CD4+, and CD8+). A 100-μL whole blood sample was incubated with 100 μL of IQ lyse (IQ Products, The Netherlands) for 10 min at room temperature. Samples were washed with PBS containing fetal bovine serum (2% vol/vol). Monoclonal mouse anti-guinea pig CD4+, CD8+, and CD3+ antibodies conjugated to R-phycoerythin (PE), fluorescein isothiocyanate, and allophycocyanin (APC) (AbD Serotec Inc., Raleigh, NC), respectively, were added to each sample preparation in series. The sample was incubated with each antibody for 10 min at room temperature in the dark, and washed before subsequent antibody incubation. Working concentrations of antibodies were according to the manufacturer's recommendations. Coulter Flow Count (100 μL) (Beckman Coulter Inc., Hialeah, FL) and PBS containing 2% fetal bovine serum (900 μL) were added to cells immediately prior to flow cytometry analysis. Samples incubated with a single or no antibody preparation served as negative controls. Samples were analyzed in a Coulter Cytomic FC500 Flow Cytometer (Becton-Dickinson, Durham, NC).
Intragastric L. monocytogenes challenge
All guinea pigs were challenged on 3 consecutive days with a 24-h interval. Each animal received approximately 1.0×104 CFU of the L. monocytogenes serotype 4b cocktail each day using a 38-cm nasogastric feeding tube (Jorgensen Laboratories Inc., Loveland, CO) fitted onto a 3-mL syringe. Animals were observed at minimum twice daily for signs of discomfort (lethargy, loss of appetite, ruffled hair coat, diarrhea).
Organ collection and microbiology
Guinea pigs were euthanized on day 4, 6, 8, 11, and 13 postchallenge using sodium pentobarbital at 100 mg/kg of body weight; liver and spleen were collected aseptically in toto. Organ samples were weighed and placed into sterile plastic tubes to which a volume equal to the weight of the sample of cold PBS containing 0.001% triton X was added. The samples were then homogenized for 30 to 60 s on ice using a single-speed tissue homogenizer (Polytrons, Kinematica, Luzern, Switzerland). A 100-μL volume was spread plated onto Rapid L'mono plate (Bio-Rad Laboratories, Inc., Richmond, CA) in duplicate and plates were incubated at 37°C for 24–48 h. L. monocytogenes colonies were counted and numbers were expressed as mean log CFU/g of organ. Remaining organ samples were stored at 4°C. When no L. monocytogenes was detected by direct plating, organ samples were subjected to microbiological enrichment procedures.
Enrichment of samples
All organ samples were subjected to enrichment procedures. An amount of 2X BHI broth (1 mL for spleen samples, 10 mL for liver samples, based on average weight of organs) was added to the organ samples, and samples were incubated at 37°C for 24–48 h. A (100 μL) aliquot was plated onto Rapid L'mono plate and Modified Oxford agar (BD Difco, Franklin Lakes, NJ) plates in duplicate and the plates were incubated at 37°C for 24–72 h to detect the presence of L. monocytogenes.
Statistical analysis
Microbiological data were analyzed using the Z-test for proportion (independent groups) (Meilgaard et al., 2006). All T-cell population data were analyzed using Microsoft Excel (Microsoft Corp., Redmond, WA) and SAS (SAS Institute, Inc., Cary, NC). The significant differences between days of treatment were determined by SAS Duncan's multiple range test, general linear model procedure. Statistical comparison of the prechallenge period (wk 0–wk 5) was based on wk 0; postchallenge period (D4–D13) was based on wk 5, p<0.05. Data were presented as means±standard deviation. The level of statistical significance for all comparisons was set at p<0.05.
Results
Listerial infection following consecutive oral challenge with 104 CFU L. monocytogenes
All animals were intragastrically challenged with approximately 1.02×104 CFU L. monocytogenes for 3 consecutive days, and infection was determined based on microbiological culture of liver and spleen samples. Diarrhea was sporadically observed and some animals exhibited signs of discomfort by D4. Two animals died, one on D8 and one on D10, although they did not exhibit persistent signs of discomfort (loss of appetite, ruffled hair coat, diarrhea). The two dead guinea pigs were negative for listerial infection based on culture enrichment of liver and spleen samples. The exact cause of death was not determined. A total of 30 animals (not including the animals that died) were included in the microbiological results. L. monocytogenes was not detected in liver or spleen of any animals by direct plating (data not shown). All organ samples were processed for microbiological enrichment (Table 1). Approximately 63% of animals became infected. All animals were infected at D4; the number of infected animals declined by D6 (p<0.05) and then increased through D13.
Different letters represent significant difference of total numbers of animal infected, p<0.05.
Liver and spleen samples were obtained from two independent experiments before oral gavage with L. monocytogenes and on days 4, 6, 8, 11, and 13 postchallenge. All organ samples were negative for L. monocytogenes by direct plating. Total number of animals infected was based on the presence of L. monocytogenes in the liver or spleen following subculture.
Effect of multiple exposure to L. monocytogenes on levels of T cells in aged guinea pigs
To gauge animal response to L. monocytogenes challenge, immune system changes in the total (CD3+), helper (CD4+) and cytotoxic (CD8+) T-cell populations were determined. During the prechallenge period, no significant change (p>0.05) in CD3+, CD4+, and CD8+ T-cell population was observed (Figs. 1 and 2), except that CD3+ T-cell populations changed significantly (p<0.05) on wk 2 and wk 3. Levels of CD3+ T cells decreased significantly (p<0.05) during the postchallenge period. CD4+ T-cell levels decreased significantly (p<0.05) on D4 and increased significantly on D6 postchallenge (Fig. 2A). CD8+ T cells increased significantly on D6 and D11 (p<0.05) (Fig. 2B).
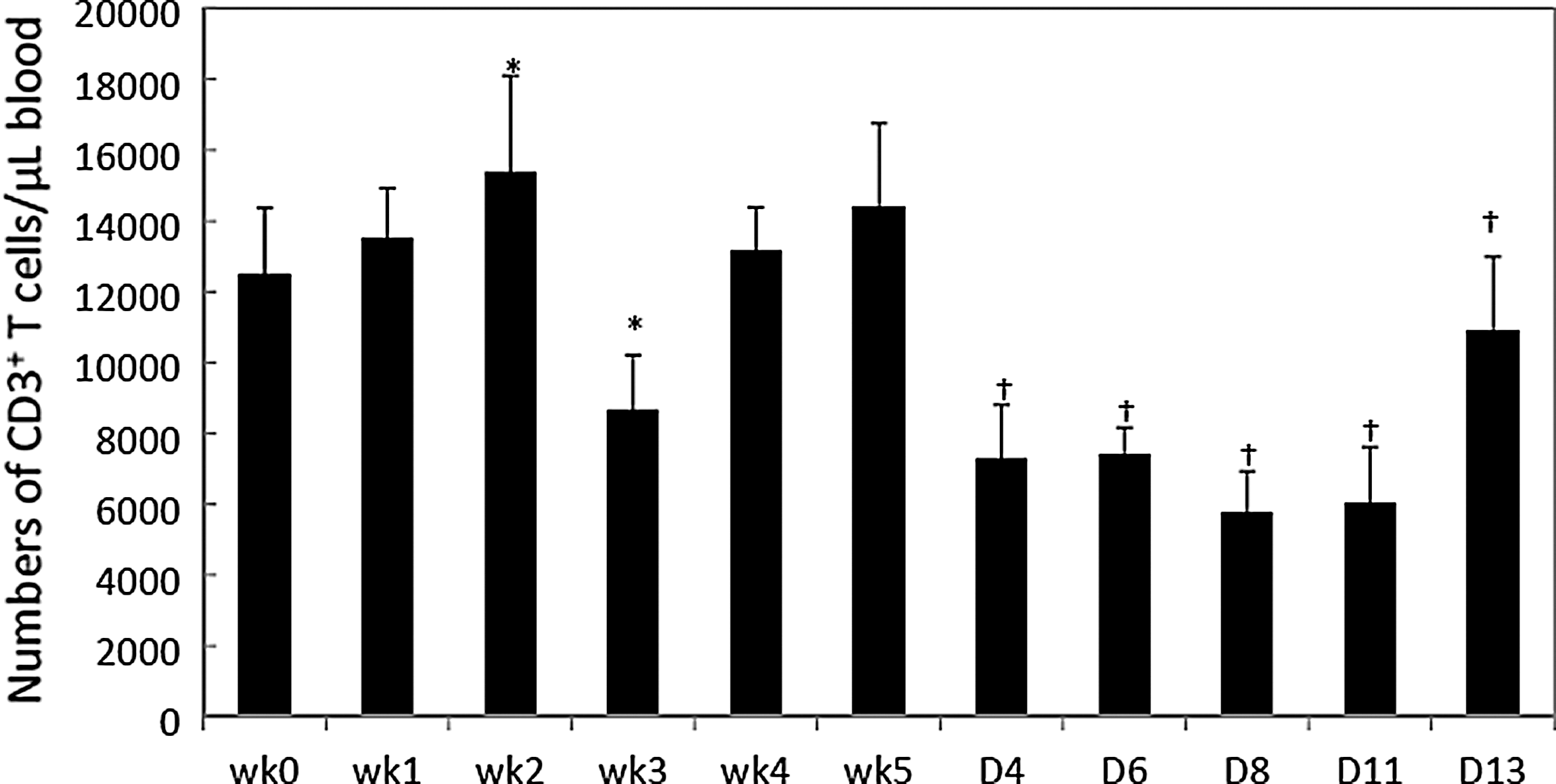
Total T (CD3+)-cell population in the prechallenge and postchallenge periods. The CD3+ T-cell levels are presented as the absolute number (cells/microliter of blood) of CD3+ T cells. The data are presented as mean±standard deviation. *Statistical comparison of prechallenge period (wk 0–wk 5) was based on wk 0, p<0.05. †Statistical comparison of postchallenge period (D4–D13) was based on wk 5, p<0.05.
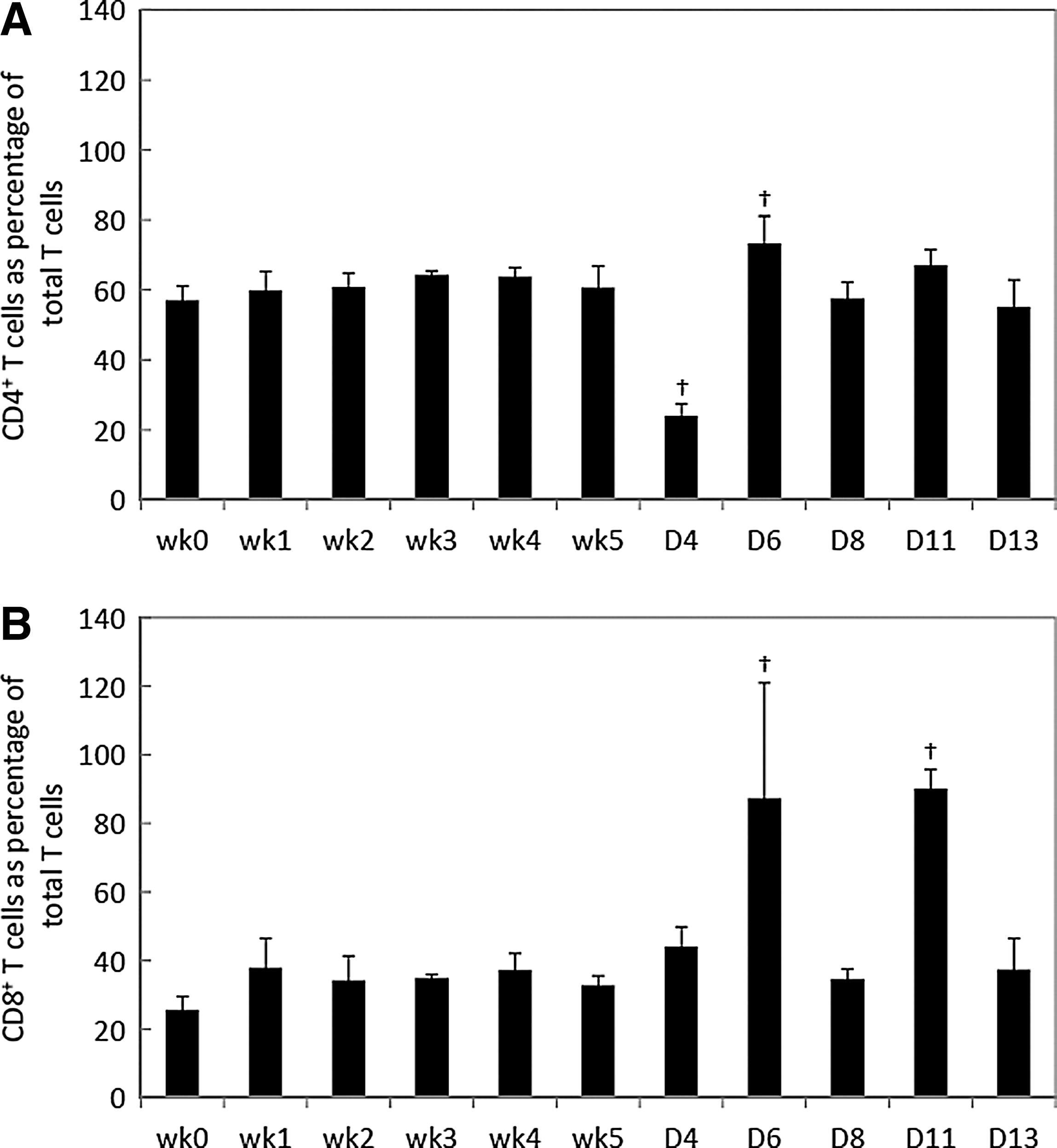
Percent CD4+ (helper) and CD8+ (cytotoxicity) T-cell prechallenge and postchallenge periods. Percent of CD4+ T cells
Discussion
Susceptibility to listerial infection is influenced by a number of factors including levels of L. monocytogenes in the food, amount of the contaminated food consumed, single or prolonged exposure, and susceptibility of the host (Maijala et al., 2001; Donnelly, 2001; McLauchlin et al., 2004; Rocourt and Cossart, 2007). On the basis of levels of L. monocytogenes and amounts of contaminated food (butter) consumed during an outbreak among immunocompromised and hospitalized patients in Finland, researchers concluded that listeriosis cases were associated with prolonged exposure to low levels (104–105 CFU/day) of L. monocytogenes in the relatively small amounts (2–28 g butter/day) of butter consumed (Maijala et al., 2001). The elderly are considered an at-risk group, and it is well established that the function of the immune system declines with age (Jackola et al., 1994; Miller, 1996; Grubeck-Loebenstein, 1997; Wick and Grubeck-Loebenstein, 1997; Ginaldi et al., 2001; Centers for Disease Control and Prevention, 2004). In the present study, retired breeder guinea pigs were used as surrogates to represent an aged human population (Pang et al., 2007; Wu et al., 2012). Age of the animals was used as the exclusive biomarker, since biomarkers for guinea pigs have not been defined (Freitas, 2003; Takizawa et al., 2004). Changes in T-cell population during the course of the study were used to gauge the ability of the aged animals to mount an immune response following repeated oral challenge with L. monocytogenes.
The 104 CFU dose used in this study was selected based on previous studies using guinea pigs demonstrating 100%, about 90%, 25%, and 13–50% of animals becoming infected when challenged orally with 108, 106, 104, and 102 CFU L. monocytogenes, respectively (Pang et al., 2007; Williams et al., 2007; Wu et al., 2012). In each of those studies, a single oral dose was given to each animal. All animals would have been considered in the “susceptible population” group since they were either pregnant or aged. Based on risk assessment data, in the United States, approximately 25 cases per year of listeriosis may occur in a susceptible population ingesting 104 CFU/g of L. monocytogenes (SOPBH, 2007). In the present study, animals were challenged with L. monocytogenes on consecutive days to represent prolonged daily consumption of a food that may typically be used for a longer time (luncheon meats, cheese, butter). In the U.S. cantaloupe outbreak, case patients may have consumed prepared cantaloupe on consecutive days, since cantaloupes weigh approximately 1.5–4 kg and a typical serving size is about 160 g. This may have contributed to the high number of cases and high mortality rate.
In this study, 63% of all animals became infected, although enrichment of the samples was required for detection of L. monocytogenes (Table 1). All animals were infected on day 4 postchallenge. The number of infected animals declined by day 6 to 14% (1 of 7) and then increased to 83% (5 of 6) by day 13 postchallenge. Studies in which animals were followed for a shorter time frame (4–8 days) show that the pathogen was cleared, or only one or two animals remained infected (Corbin and Harty, 2004; Pang et al., 2007; Wu et al., 2012). Using an enrichment protocol, we demonstrated that Listeria may not be cleared and can persist for an extended period after repeated challenge. The initial immune response by the aged guinea pigs was sufficient to combat the infection during the first few days following challenge, but was not robust enough to clear the infection (Table 1).
T cells have been shown to play an important role to further confront listerial infection after the initial control of L. monocytogenes by the innate immune response. Studies demonstrated that notable changes in the CD8+ T-cell response peaks at 7–9 days postinfection (Busch et al., 1998; Pope et al., 2001; Harty and Badovinac, 2002; Badovinac et al., 2002). In the present study, L. monocytogenes triggered strong CD8+ T-cell increase by D6 postchallenge, indicating that cytotoxic T cells were produced to clear L. monocytogenes. The consecutive listerial challenges resulted in a second increase in the CD8+ T-cell population on D11. The effect of successive L. monocytogenes challenges resulted in a decrease of the total T-cell population. The decline was consistent with the persistence of listerial infection till D13. The T cells were utilized either for clearance of the pathogen or production of memory T cells.
CD4+ T cells also play a protective role in control of listerial infection. CD4 T cells assist in pathogen recognition by CD8 T cells (Mittrucker et al., 2000; Kursar et al., 2002; Shedlock and Shen, 2003). In the present study, CD4+ T-cell populations decreased on D4 and then increased on D6 (p<0.05). A previous study conducted in our lab (Wu et al., 2012) showed that CD4+ T cells underwent an immediate decline on D3 postchallenge; levels of CD4+ T cells then increased from D6 to D20 postchallenge, although levels of CD4+ T cells remained lower than wk 3 (before challenge). In the present study, CD4+ T cells increased significantly on D6 and remained higher than wk 5 (before challenge) throughout the 13-day experiment. The differences between the studies may be associated with the use of a single lower challenge inoculum (100 CFU), which resulted in lower CD4+ T-cell responses than observed in the present study. Corbin and Harty (2004) indicated that the duration of infection influenced the magnitude of antigen-specific CD4+ and CD8+ T-cell responses.
Conclusion
Results of the present study demonstrated that repeated daily exposure to a low dose of L. monocytogenes can result in infection of long duration (≥13 d) or delayed onset. Studies conducted in pregnant guinea pigs using a single high-dose (≥106 CFU) challenge showed that infection was rarely detected beyond day 6 postchallenge (Williams et al., 2007). The guinea pigs mounted an immunological response typical for an acute infection with an intracellular pathogen such as L. monocytogenes. Prolonged daily exposure to low levels of L. monocytogenes may result in long-term depression of T cells, which may potentially overwhelm the ability of an aged animal to combat the pathogen, resulting in infection and perhaps more serious sequelae.
Footnotes
Acknowledgments
We thank Dr. Elizabeth Dodemaide for performing oral gavage on guinea pigs. The work was supported by a research grant from the Center for Advanced Food Technology, Rutgers, The State University of New Jersey, New Brunswick, New Jersey.
Disclosure Statement
No competing financial interests exist.