
Abstract
Dairy cattle serve as a potential source for Campylobacter infection in humans. Outbreaks associated with consumption of either Campylobacter contaminated raw milk or contaminated milk after treatment were previously recorded in the United States. Further, starlings have been implicated in the spread of bacterial pathogens among livestock. Here, we determined the prevalence, genotypic, and phenotypic properties of Campylobacter isolated from fecal samples of dairy cattle and starlings found on the same establishment in northeastern Ohio. Campylobacter were detected in 83 (36.6%) and 57 (50.4%) out of 227 dairy and 113 starling fecal samples, respectively. Specifically, 79 C. jejuni, five C. coli, and two other Campylobacter spp. were isolated from dairy feces, while all isolates from starlings (n=57) were C. jejuni. Our results showed that the prevalence of C. jejuni in birds was significantly (p<0.01) higher than that in dairy cattle. The pulsed-field gel electrophoresis analysis showed that C. jejuni were genotypically diverse and host restricted; however, there were several shared genotypes between dairy cattle and starling isolates. Likewise, many shared clonal complexes (CC) between dairy cattle and starlings were observed by multilocus sequence typing (MLST) analysis. As in humans, both in cattle and starlings, the CC 45 and CC 21 were the most frequently represented CCs. As previously reported, CC 177 and CC 682 were restricted to the bird isolates, while CC 42 was restricted to dairy cattle isolates. Further, two new sequence types (STs) were detected in C. jejuni from dairy cattle. Interestingly, cattle and starling C. jejuni showed high resistance to multiple antimicrobials, including ciprofloxacin, erythromycin, and gentamicin. In conclusion, our results highlight starlings as potential reservoirs for C. jejuni, and they may play an important role in the epidemiology of clinically important C. jejuni in dairy population.
Introduction
C
Wild birds may play a role in the epidemiology of human infection by indirectly transmitting Campylobacter among livestock (Jones, 2001; Rosef et al., 1983; Cabrita et al., 1992; Waldenström et al., 2005). Campylobacter could spread from farms to the environment and could further spread to environmental reservoirs such as wild birds, which may facilitate their transmission from one farm to another (Waldenström et al., 2005). In a study in northwest England using multilocus sequence typing (MLST), most commonly recovered genotypes from dairy cattle, wildlife, and environmental sources were frequently associated with human disease (Kwan et al., 2008b). Further, wild European Starlings (Sturnus vulgaris) were suggested to be a potential source of human and livestock infection, because of their high shedding of Campylobacter (Colles et al., 2008; Nielsen et al., 2004) as well as their capability to disseminate infections between farms (LeJeune et al., 2008). Furthermore, starlings have the tendency to invade and gather in thousands at dairy farms and animal feeding operations (Linz et al., 2007).
Here, we investigated the role that starlings play as a source of Campylobacter infection and/or transmission to dairy cattle through contaminated droppings. A cross-sectional study was conducted to determine the prevalence, genotypic relatedness between Campylobacter isolated from dairy cattle and starlings captured on those farms in northeastern Ohio. Our results indicated that starlings carry C. jejuni genotypes that are indistinguishable from those exist in cattle and thus play an important role in the epidemiology of clinically important C. jejuni in dairy population.
Materials and Methods
Collection and processing of fecal samples from dairy cattle and starlings
We investigated the occurrence of C. jejuni and C. coli in dairy cattle and starlings in dairy establishments in northeastern Ohio. A total of 227 fresh dairy fecal samples as well as 113 starling fecal samples were collected during the summer and early fall of 2009 from 11 dairy farms (Table 1). The sampling time marks the period of peak bovine shedding of Campylobacter and when starlings congregate in large flocks on farms. Ten grams of cattle feces were collected, and starlings were captured during each visit using mist nets or decoy cages (Table 1). Starlings were euthanized by cervical dislocation, and carcasses were transported to the laboratory on ice.
A total of 86 Campylobacter isolates occurred in 83 dairy fecal samples, including two Campylobacter spp. other than C. jejuni and C. coli.
Starlings could not be captured.
Isolation and identification of Campylobacter spp. from fecal samples
One gram of each fecal sample was enriched in Preston broth for 48 h at 42°C microaerobically (Krause et al., 2006). From the enrichments, 100 μL was spread onto modified Cefoperazone Charcoal Deoxycholate Agar (mCCDA) plates and incubated for 48 h at 42°C microaerobically (Engberg et al., 2000). Three to five colonies suspected as Campylobacter were sub-cultured onto Muller-Hinton (MH) agar. Genomic DNA was isolated using DNA Purification Kit (Epicenter Biotechnologies, Madison, WI), and identity was confirmed by polymerase chain reaction (PCR). The PCR analysis targeted a 16S rRNA gene, mapA, and ceuE, which indicated the specific detection of Campylobacter spp., C. jejuni, and C. coli, respectively (Denis et al., 1999; Linton et al., 1997). The C. jejuni 81-176, originally isolated from an outbreak associated with raw milk (Bacon et al., 2000), and C. coli (ATCC 33559) strains were used as positive controls. PCR reactions with no DNA templates were used as negative control.
Pulsed Field Gel Electrophoresis (PFGE)
The PFGE analysis was performed as described previously (Ribot et al., 2001; Sanad et al., 2011). Briefly, optical density (OD)–adjusted culture suspensions were gently mixed with 1% SeaKem Gold agarose (Fisher Scientific, Fair Lawn, NJ) to prepare agarose plugs. The plugs were incubated with shaking (200 rpm) in lysis buffer (50 mM Tris, 50 mM EDTA [pH 8.0]), 1% sarcosine, 0.1 mg mL−1 of proteinase K) for 1 h at 55°C, washed 4×, and suspended in TE. Plugs were sliced and digested overnight with SmaI at room temperature. The DNA fragments in the digested slices were separated by electrophoresis for 20 h using the CHEF Mapper system (Bio-Rad, Hercules, CA) with initial and final switch times of 6.75 and 38.35 s, respectively, gradient of 6 V/cm, and an angle of 120°. The resulting PFGE patterns were analyzed using the BioNumerics 5.1 software (Applied Maths Inc., Austin, TX). Similarity and clustering analysis of the PFGE patterns were performed using the Dice Coefficient and the unweighted pair-group method with arithmetic averages (UPGMA) with optimization of 1% and position tolerance of 1.5% (Ribot et al., 2001). Lambda Ladder PFG Marker −50-1,000 kb (New England BioLabs, Ipswich, MA) was used as a molecular marker.
Antimicrobial resistance (AR) typing
Minimum inhibitory concentration (MIC) of C. jejuni and C. coli isolates were determined using Sensititre Campy plates (TREK Diagnostic Systems Inc., Cleveland, OH). A total of 117 C. jejuni isolates (72 dairy cattle and 45 starlings) representing different PFGE clusters were analyzed. Isolates un-typeable by PFGE were not included for analysis. The AR typing was performed as described previously (Sanad et al., 2011). The susceptibility and resistance breakpoints were interpreted according to the Clinical and Laboratory Standards Institute (CLSI, 2006). C. jejuni 81-176 was used for quality control.
MLST
The MLST analysis was performed as described previously (Dingle et al., 2001; Sanad et al., 2011) on a total of 75 C. jejuni isolates (44 dairy cattle and 31 starling) that represented the main PFGE clusters and different locations (farms). Briefly, loci from seven housekeeping genes were amplified by PCR, and PCR products were treated with ExoSAP-IT (Affymetrix, Inc., Cleveland OH) and sequenced in both directions. The sequences were aligned using ClustalW (
Statistical analysis
The adjusted sample size was calculated using a hypothetical prevalence in wild birds of 5%. Thus, initial estimates for simple random sampling was as follows: study power, 0.7; α=0.05; and error margin, 0.1. For adjusted clustering, we used STATA, version, 9.0 and interclass correlation coefficient=0.2. A Chi square test was used to evaluate prevalence data. A p-value of <0.05 was chosen for statistical significance.
Results
Occurrence and distribution of Campylobacter spp. in dairy cattle and starling feces
Campylobacter were detected in 83/227 (36.6%) dairy fecal samples and 57/113 (50.4%) starling fecal samples. A total of 86 Campylobacter isolates were recovered from 83 fecal samples. C. jejuni represented the majority of the isolates (n=79, 91.9%) followed by C. coli (n=5, 5.8%), while two isolates (2.3%) were species other than C. jejuni and C. coli (Table 1). A total of 57 C. jejuni and no C. coli were isolated from starling feces. Our results showed that the prevalence of C. jejuni in starlings was significantly (p<0.01) higher than that in dairy cattle.
C. jejuni strains were genotypically diverse among different host species
The PFGE was successfully performed on 130 of the 136 dairy cattle and starling C. jejuni isolates, while six isolates could not be typed. These six isolates were also untypeable when digested with KpnI or using formaldehyde (data not shown). PFGE profiles indicated highly diverse genotypes, especially among isolates from different farms (Fig. 1). Four main clusters identified with cluster I which mainly consisted of cattle C. jejuni (Fig. 1). All clusters included profiles of C. jejuni from both dairy cattle and starlings with different percentages of similarities. However, C. jejuni isolates from dairy cattle and starlings with 100% similarity were observed in 11 subclusters (see arrows in Fig. 1), and three of those subclusters possessed shared macrorestriction profiles (MRPs) for C. jejuni in both host species from the same farm location (see stars in Fig. 1). The other eight subclusters constituted cattle and starling C. jejuni from different farms including one with 5 isolates from 4 different farms (see asterisk Fig. 1). None of cattle C. jejuni isolates from three farms (A, B, C-m) where no starlings were captured shared genotypes with C. jejuni from starlings at the other eight farms (Table 1 and Fig. 1).

Dendrogram showing the pulsed-field gel electrophoresis patterns for SmaI restricted dairy cattle and starlings Campylobacter jejuni. Numbers on bootstraps represent cophenetic correlations. Arrows indicate the shared (100% similarity) macro restriction profiles (MRPs) of C. jejuni isolates from dairy cattle and starlings. Stars represent the shared (100% similarity) MRPs from both hosts within the same location (farm). Asterisk shows the shared (100% similarity) MRPs from both hosts but obtained from different locations (farms). A–J, farms; D, dairy; Br, birds.
Antimicrobial susceptibility of dairy and starling C. jejuni
C. jejuni from dairy cattle were resistant to several antimicrobials including, ciprofloxacin, erythromycin, gentamicin, tetracycline, and clindamycin (Fig. 2). Resistance to tetracycline was most common among dairy cattle C. jejuni (68.0%) (Fig. 2). Furthermore, 28 (38.9%) C. jejuni isolates were resistant only to tetracycline, 3 (4.16%) were resistant only to two antimicrobials, 7 (9.7%) were resistant to only 3 antimicrobials, while 18 (25%) were resistant to 4 or more antimicrobials including ciprofloxacin, tetracycline, nalidixic acid, erythromycin, and clindamycin (Fig. 2). Fifteen isolates (20.8%) were pan-susceptible.
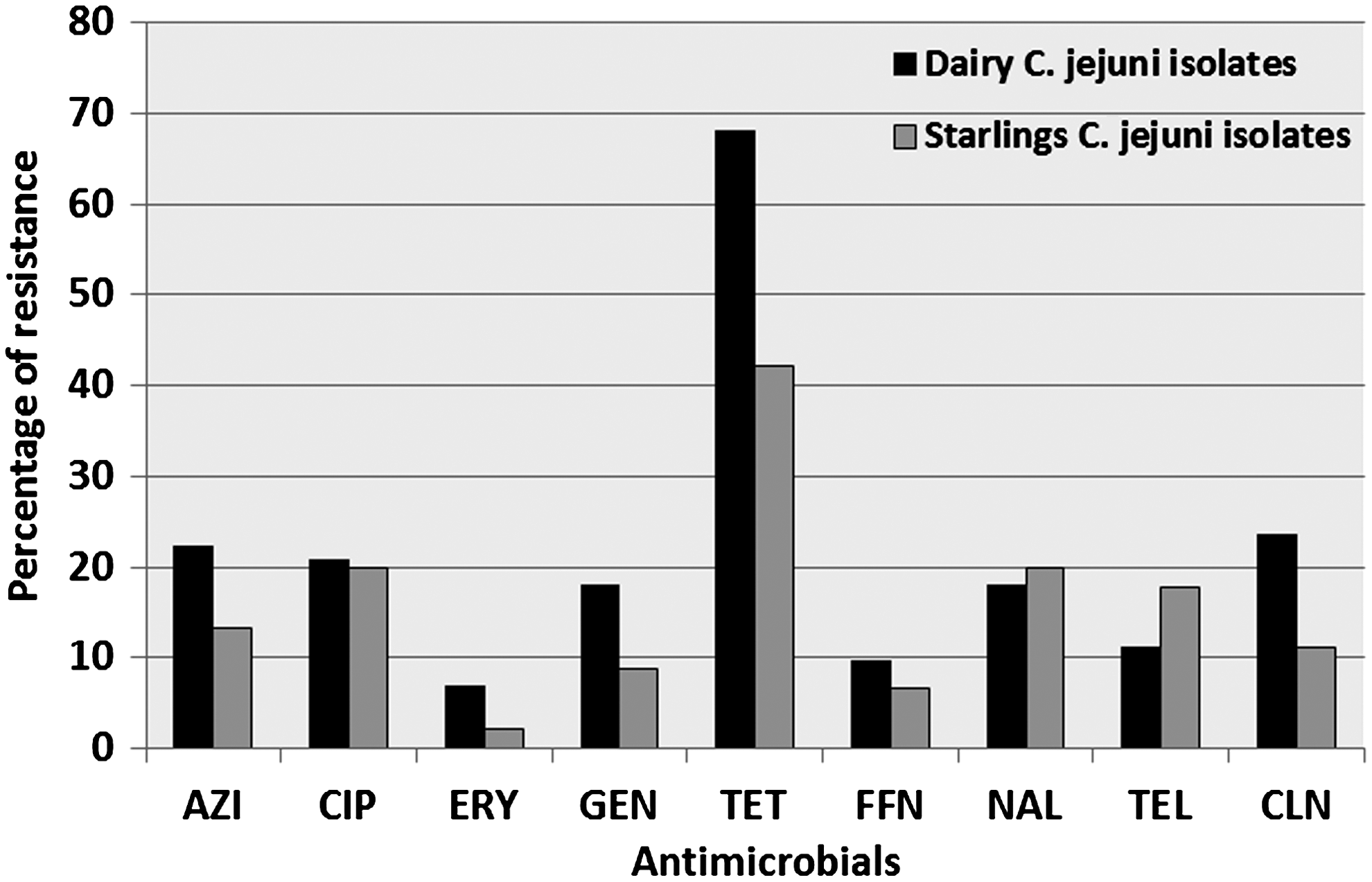
Antimicrobial resistance of Campylobacter jejuni isolated from dairy cattle (n=72) and starlings (n=45). Percentage of total isolates resistance to different antimicrobials used in this study is shown.
Although the resistance of cattle and starling C. jejuni were different for most antimicrobials, the starling C. jejuni also exhibited resistance to several antimicrobials including ciprofloxacin, gentamicin, nalidixic acid, tetracycline, and clindamycin. Resistance to tetracycline was also common among bird C. jejuni isolates (42.2%), while only one (2.2%) isolate showed resistance to erythromycin (Fig. 2). Of the 45 bird isolates, 8 (17.8%) were resistant only to tetracycline, 5 (11.1%) were resistant to two antimicrobials, 7 (15.5%), were resistant only to three antimicrobials, 5 (11.1%) were resistant to four or more antimicrobials, and 19 (42.2%) were pan-susceptible (Figs. 2 and 3). The control C. jejuni 81-176 strain is pTet plasmid cured (Rajashekara et al., 2009) and showed resistance only to nalidixic acid.
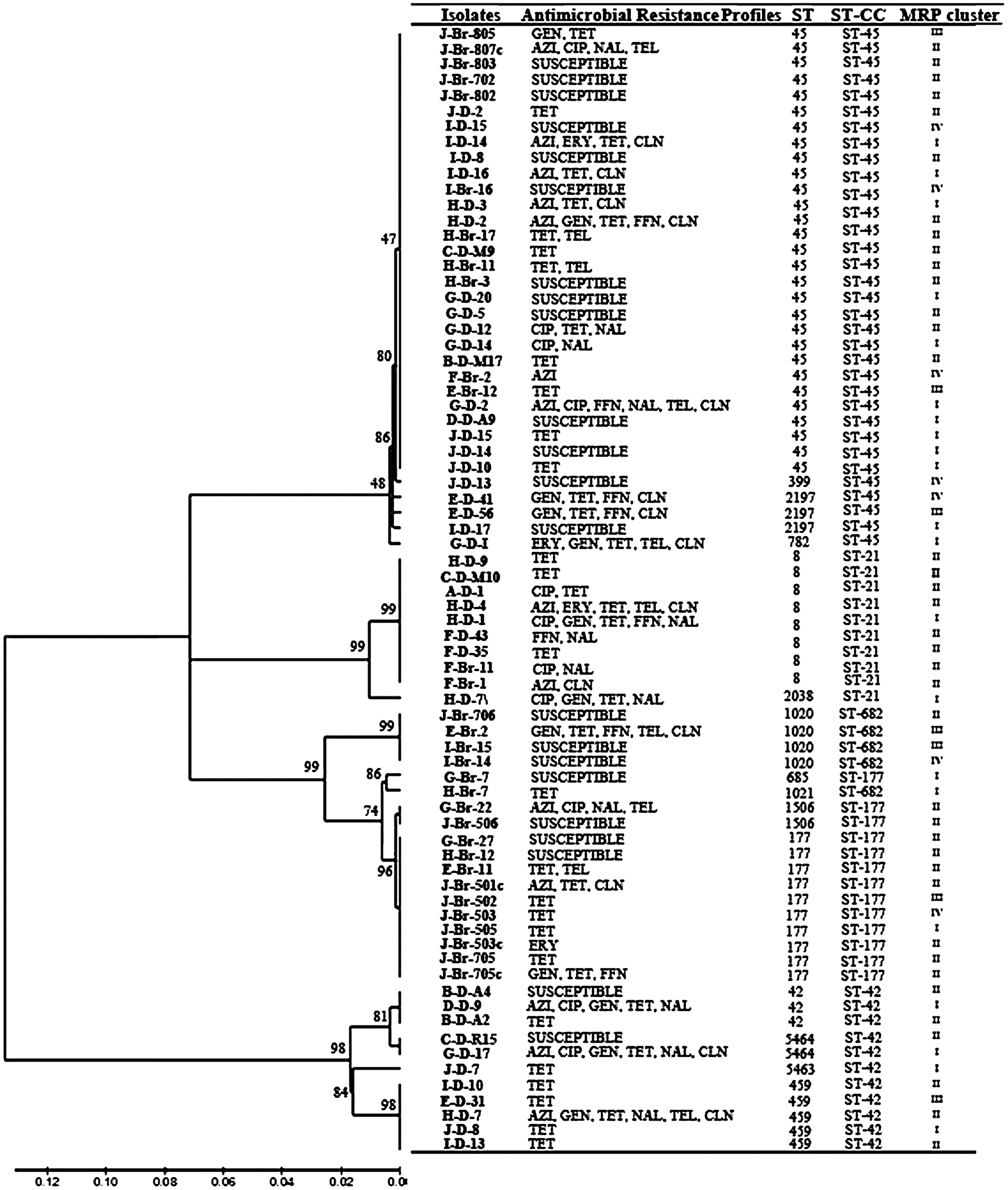
Multilocus sequence typing (MLST) analysis and antimicrobial resistance profiles of Campylobacter jejuni isolates from dairy cattle and starlings. The percentage values of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown above the branches. The tree is drawn to scale, with branch lengths (above the branches) in the same units as those of the evolutionary distances used to infer the phylogenetic tree. Antimicrobial resistance profiles, sequence types, and macro restriction profile (MRP) clusters of dairy cattle and starlings C. jejuni isolates were combined to the tree. A–J, visited farms; D, dairy; Br, birds.
MLST identified STs belonging to clonal complexes of public health significance
In both species, a total of 15 different STs (eight dairy cattle and five starlings) were identified, and two STs (ST-45 and ST-8) were shared between both species (Fig. 3). Specifically, 34 (23 dairy cattle and 11 starling) isolates, representing four STs, belonged to CC 45. An additional nine isolates (seven dairy cattle and two starlings) were grouped into ST-8 that belonged to CC 21 (Fig. 3). However, 11 dairy cattle C. jejuni representing four STs, including the two newly identified STs (ST-459, ST-42; novel STs 5463 and 5464), belonged to CC 42. In addition, three isolates were identified as ST-2197, one as ST-399, and one as ST-782, which all belonged to CC 45. One isolate was identified as ST-2038, which belonged to CC 21. On the other hand, 10 starlings isolates were identified as ST-177, two as ST-1506, and one as ST-685, which all belonged to CC 177. Further, four isolates were identified as ST-1020 and one as ST-1021, which belonged to CC 682 (Fig. 3). The complete profile for two isolates could not be obtained.
Discussion
Although the association of Campylobacter spp. with dairy cattle in the United States has been investigated previously, available literature reported variable prevalence. Our dairy cattle–associated Campylobacter data (36.6%; Table 1) is within the range of 0–51.2% previously reported in the United States (Harvey et al., 2004; Wesley et al., 2000; Sato et al., 2004; Bae et al., 2005; Dodson and LeJeune, 2005; Englen et al., 2007), as well as in concordance with those ranges reported outside the United States (35.9%) (Kwan et al., 2008a). The prevalence data for Campylobacter in wild birds in the United States are very limited. For instance, C. jejuni was isolated from six avian families with the highest prevalence of 25% (Keller et al., 2011). The occurrence of Campylobacter spp. in starlings in our study was higher (50.4%) than in the previous reports from other countries such as Sweden, 21.6% (Waldenström et al., 2002), and New Zealand, 30.6% (French et al., 2009). However, prevalence of 2–50% in wild bird species, including starlings in England, has been reported (Hughes et al., 2009). Similar to previous studies, C. jejuni represented all the retrieved isolates from starlings (Luechtefeld et al., 1980; Kapperud and Rosef, 1983; Waldenstrom et al., 2002; Broman et al., 2004; Keller et al., 2011), as well as the majority of dairy cattle fecal samples (Table 1) (Wesley et al., 2000; Bae et al., 2005). Our prevalence data provide evidence that dairy cattle and starlings constitute significant reservoirs for these pathogens, promoting their persistence in dairy cattle–derived products and environment, which might pose a risk for humans through consumption of dairy or beef products.
Remarkably, PFGE analysis revealed profiles of some C. jejuni isolates possessing genotypes that were 100% similar on 29 different occasions (Fig. 1), 11 of them including shared isolates from both species (see arrows in Fig. 1). These observations support the findings of studies conducted in cattle in other countries (Oporto et al., 2007; Grove-White et al., 2010; Litrup et al., 2007; de Haan et al., 2010; Açik and Çetinkaya, 2006) as well as in wild birds (Broman et al. 2004). Additionally, we found three instances where C. jejuni isolates from both host species, originating from the same locations, clustered together with isolates originating from different farms, indicating potential dissemination of strains by starlings (see stars in Fig. 1). These results support the notion that starlings, which are highly mobile (Johnson and Glahn, 1992), can serve as a vector for transmission of pathogens between farms (LeJeune et al., 2008).
Campylobacter resistance to antibiotics is an increasingly important emerging threat to public health (Châtre et al., 2010; Luangtongkum et al., 2009; Englen et al., 2005). Previous studies have demonstrated that cattle-associated Campylobacter can resist antimicrobial drugs of clinical importance to human health. For example, 47.4%, 4%, and 2.5% of C. jejuni collected from dairy cows in the United States were resistant to tetracycline, nalidixic acid, and ciprofloxacin, respectively (Englen et al., 2007). Additionally, 47.7% and 49.1% of C. jejuni from dairy and feedlot cattle, respectively, were resistant to tetracycline (Englen et al., 2005, 2007). In agreement with these findings, our AR analysis showed that a high percentage of the C. jejuni from dairy cattle were resistant to multiple antimicrobials (Figs. 2 and 3). In our study, approximately 21% and 7% of the cattle C. jejuni were resistant to ciprofloxacin and erythromycin, respectively (Fig. 2). This is an important difference compared to previous studies in the United States that reported a relatively lower frequency of resistance to ciprofloxacin (1.8–5%) and erythromycin (0.4–2.9%) in C. jejuni isolated from different cattle operations (Bae et al., 2005; Englen et al., 2005, 2007). Use of antimicrobials in the dairy cattle and other livestock has been attributed to an increase in AR among pathogens (Oliver et al., 2011). Our data support the predictions that livestock-associated Campylobacter are becoming increasingly resistant to important antibiotics (Châtre et al., 2010; Luangtongkum et al., 2009; Oliver et al., 2011). Similarly, starling-associated C. jejuni also displayed higher resistance (11.1%) than those previously reported outside the United States from wild birds. For example, in a previous study, the resistance rate of bird isolates for several antimicrobials was 0.7–3.6% (Waldenström et al., 2005). Horizontal transfer of resistance features among Campylobacter species as well as transmission of resistant Campylobacter between different species may provide potential mechanisms for emerging AR (Velazquez et al., 1995; Sundlof, 2000). These observations highlight the need for strict surveillance of antimicrobials used in dairy cattle operations in order to facilitate interventions and curb further emergence of antibiotic-resistant Campylobacter.
The MLST analysis confirmed that dairy cattle and starling C. jejuni were diverse and certain C. jejuni may be highly host associated (CC 42, cattle; CC 177 and CC 682, starlings [Waldenstrom et al., 2002; Colegrave and Buckling, 2005]). In fact, CC 21, CC 45, CC 42, and CC 61 were also predominant in cattle samples from other countries (de Haan et al., 2010; Dingle et al., 2002; Kwan et al., 2008a,b; Levesque et al., 2008). The detection of CC 45, CC 21, and CC 42 (Fig. 3) in this study is of particular interest, since these complexes constitute three of six CCs that represent 60% of C. jejuni associated with human disease (Dingle et al., 2001; Grove-White et al., 2010). The clear absence of unique starling-associated C. jejuni strains (ST-177 and ST-685) in dairy cattle may suggest that (i) infection is likely transmitting from dairy cattle to birds (Hughes et al., 2009) and starlings are involved in transmission of pathogen between dairy operations, and (ii) some C. jejuni strains may be host adapted and survive (transmit) poorly in disparate host reservoirs.
No specific relationship was observed between MRPs, STs, and AR (Fig. 3). PFGE is typically more discriminatory than the MLST (Fakhr et al., 2005); further, the low GC content and the ability for natural competence makes C. jejuni weakly clonal (Wassenaar and Newell, 2000; Dingle et al., 2001). Despite the very weak clonal nature, the identification of indistinguishable MRPs among C. jejuni from dairy cattle and starlings clearly suggests a role for starlings in the epidemiology of this pathogen in dairy production.
Taken together, our studies indicate starlings play a role in transmission of Campylobacter in dairy cattle populations as well as dissemination of C. jejuni that are commonly associated with human infections. The potential of dairy cattle and starling Campylobacter to resist multiple antimicrobials, including those antibiotics of choice for treatment of human cases, reassert the public health significance of Campylobacter infections in humans through consumption of contaminated dairy and dairy products. Thus, more molecular epidemiological studies would enhance the efforts to establish effective control and preventive measures, and limit the public health impact of this pathogen from dairy cattle and associated reservoirs.
Footnotes
Acknowledgments
We thank Dr. Wondwossen Gebreyes for the critical review of the manuscript. Research in Dr. Rajashekara's laboratory is supported by the Ohio Agricultural Research and Development Center (OARDC), Ohio State University, and the Agriculture and Food Research Initiative (AFRI) grant #2012-68003-19679, U.S. Department of Agriculture. This research was partially supported by funds appropriated to Dr. LeJeune (NRI-2006-01227). Dr. Sanad was supported from the Egyptian Ministry of Higher Education scholarship.
Disclosure Statement
No competing financial interests exist.