
Abstract
Kudoa septempunctata is a myxosporean parasite of Paralichthys olivaceus (olive flounder) and causes a foodborne illness that affects more than 100 cases in Japan each year. We previously reported that the consumption of raw olive flounder meat containing a high concentration of K. septempunctata spores induces transient but severe diarrhea and emesis through an unknown mechanism. Here, we demonstrate that K. septempunctata sporoplasm plays an important role in mediating the toxicity of K. septempunctata. When K. septempunctata spores were inoculated in Caco-2 human intestinal cells, K. septempunctata sporoplasms were released from spores, and they invaded the cells. Electron microscopic observations revealed that the sporoplasm invasion severely damaged the Caco-2 cells. The inoculation of K. septempunctata spores eliminated the transepithelial electrical resistance (TER) across the cell monolayer. Inhibiting the invasion of the sporoplasms prevented the observed loss in cell layer integrity, as illustrated by the rapid elimination of the TER. These results suggest that the invasion by sporoplasms severely damaged individual intestinal cells, resulting in a loss of cell monolayer integrity.
Introduction
O
The K. septempunctata spore has six or seven shell valves and polar capsules (Matsukane et al., 2010). The spore of this parasite has a pouch-like shape and is about 10 μm in diameter (Matsukane et al., 2010). K. septempunctata cells dwell in the trunk muscles of P. olivaceus, and its spores are found uniformly distributed throughout P. olivaceus muscles. It has been suggested that the life cycle of K. septempunctata is maintained between P. olivaceus and an oligochaete or polychaete worm, but this has not been thoroughly studied (El Matbouli et al., 1998). Oral administration of K. septempunctata spores to suckling mice caused fluid accumulation in the gut within 1.5 h of administration, with recovery after 4 h (Kawai et al., 2012). The feeding of olive founder meat containing K. septempunctata spores to house musk shrews induced vomiting after 20–30 min with recovery within 2 h (Kawai et al., 2012). Together, these experiments demonstrate the short lag phase and quick recovery from this parasite, and the symptoms observed in the animal experiments coincide with the symptoms exhibited by human patients with this disease (Kawai et al., 2012). Despite this research, however, the mechanisms of this disease are poorly understood. Many species of protozoa induce diarrhea. In fact, protozoa such as Giardia lamblia, Toxoplasma gondii, Cryptosporidium parvum, and Entamoeba histolytica can invade and reproduce in the intestine, which is a cause of infection-associated diarrhea (Brandborg et al., 1967; Chen et al., 2000; Dubey et al., 1997; Saha et al., 1977; Yoshida et al., 2011). Since some myxosporean parasites invade and reproduce in the intestinal cells of the worm (El Matbouli et al., 1998), it has been hypothesized that Kudoa spp. are also able to invade the intestinal cell. However, it has not been shown whether K. septempunctata is able to invade and reproduce in the intestine of the worm. Moreover, we know that K. septempunctata causes disease in humans, but the behavior of K. septempunctata in the human intestine has not been well studied. In order to examine possible mechanisms by which K. septempunctata may cause diarrhea, we studied the toxicity and behavior of K. septempunctata in cultured human intestinal cells.
Materials and Methods
Cell monolayer permeability assay
The human adenocarcinoma cell line Caco-2 cells (American Type Cell Culture [ATCC] HTB37) was normally maintained in Dulbecco's modified Eagle's medium (Life Technologies, Grand Island, NY) supplemented with 10% fetal calf serum (FCS), antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin; Life Technologies) and 1% non-essential amino acids (Life Technologies) at 37
In some experiments, purified K. septempunctata spores were suspended in phosphate-buffered saline (PBS) and sonicated by Branson Sonifier 200 (Branson Ultrasonics, Danbury, CT). The sonicated spores (equivalent to 5×105 spores/well) were inoculated in cell culture inserts, and the TER was measured.
To examine toxin production, K. septempunctata spores (1×107 spores/mL) were suspended in EDM supplemented with 0.08% MITO+ Serum Extender. The suspension was incubated 37°C for 18 h. After centrifugation at 1,500×g for 15 min at 4°C, the supernatant was recovered. The culture medium of differentiated Caco-2 cells was replaced with the supernatant of K. septempunctata spores, and the TER was measured.
Sporoplasm release assay
K. septempunctata spores purified as above were suspended in PBS, H2O, or H2O (pH 2) to a final concentration of 107 spores/mL, with or without protease inhibitor (Nacalai Tesque, Inc., Kyoto, Japan). These suspensions were then incubated with FCS, trypsin (Thermo Fisher Scientific Inc., Rockford, IL), or pepsin (Wako Pure Chemical Industries, Ltd., Osaka, Japan) at the final concentration indicated in Table 1. The suspensions were incubated at room temperature for 1 h, and sporoplasm release was observed under a bright-field microscope. The release of sporoplasm was counted in 10 randomly selected microscopic fields, and a positive result was indicated by sporoplasm release from >30% of the spores.
FCS, fetal calf serum; PBS, phosphate-buffered saline.
Transmission electron microscopy (TEM)
After plating Caco-2 cells (8×105 cells/dish) in 6-cm cell culture dish coated with Cellmatrix type I collagen solution (Nitta Gelatin Inc., Osaka, Japan), the cells were differentiated as described above. Three days later, the purified K. septempunctata spores (6×106 spores/dish) were inoculated into cells, and the cultures were incubated at 37°C for 1 h. After incubation, the cultures were fixed with 2% paraformaldehyde, 2% glutaraldehyde in 0.1 M phosphate buffer (PB) pH 7.4 at 37°C, and incubated at 4°C for 30 min to lower the temperature. Thereafter, they were fixed with 2% glutaraldehyde in 0.1 M PB at 4°C overnight. Following fixation, the samples were rinsed three times with 0.1 M PB for 30 min each, followed by post fixation with 2% osmium tetroxide in 0.1 M PB at 4°C for 1 h. The samples were dehydrated through an ethanol series (50%, 70%, 90%, 100%). The schedule was as follows: 50% and 70% for 15 min each at 4°C, 90% for 5 min at room temperature, and three changes of 100% for 5 min each at room temperature. After dehydration, the samples were transferred to a resin (Quetol-812; Nisshin EM Co., Tokyo, Japan) and polymerized at 60°C for 48 h. Ultra-thin sections (70 nm) of the blocks were prepared with a diamond knife by using an ultramicrotome (Ultracut UCT; Leica Microsystems GmbH, Wetzlar, Germany), and the sections were placed on copper grids, stained with 2% uranyl acetate at room temperature for 15 min, and then rinsed with distilled water, followed by secondary staining with lead stain solution (Sigma-Aldrich Co., St. Louis, MO) at room temperature for 3 min. The grids were observed by TEM (JEM-1200EX; JEOL Ltd., Tokyo, Japan) at an acceleration voltage of 80 kV. Digital images were taken with a charge-coupled device (CCD) camera (Veleta; Olympus Soft Imaging Solutions GmbH, Münster, Germany).
Scanning electron microscopy (SEM)
Caco-2 cells (8×105 cells/dish) were seeded and differentiated as mentioned above. Three days later, the purified K. septempunctata spores (6×106 spores/dish) were inoculated into cells, and the cultures were incubated at 37°C for 1 h. After incubation, the samples were fixed and dehydrated as described above. The samples were treated with tert-butyl alcohol three times for 30 min each, followed by freezing at 4°C. The frozen samples were dried using a dry vacuum pump (DAP-6D; ULVAC KIKO, Inc., Miyazaki, Japan) with slow decompression. After drying, the samples were coated with a thin layer (30 nm) of osmium by using an osmium plasma coater (NL-OPC80NS; Nippon Laser & Electronic Laboratory, Nagoya, Japan). The samples were observed by SEM (S-800; Hitachi High-Tech Fielding Co., Tokyo, Japan) at an acceleration voltage of 10 kV.
Inhibition of sporoplasm release and confocal microscopy
After plating Caco-2 cells (4×105 cells/culture) in a cover glass coated with Cellmatrix type I collagen solution, the cells were differentiated as described above. Three days later, the purified K. septempunctata spores (3×106 spores/culture) were inoculated into Caco-2 cells with or without cytochalasin D (Wako Pure Chemicals) dissolved in DMSO (Wako Pure Chemicals), and the cultures were incubated at 37°C for 1 or 2 h. The cells were washed two times with PBS and fixed with 4% paraformaldehyde in PB for 30 min at room temperature. After fixation, the cells were washed two times with PBS, and then incubated with chicken anti–K. septempunctata antiserum in PBS containing 10% normal goat serum at 4°C for 18 h. Chicken anti–Kudoa antiserum was raised against sonicated K. septempunctata spores according to the method described by Matsuda et al. (1999). After incubation, the cells were washed five times with PBS, and then incubated with Alexa 488 goat anti–chicken IgG (Life Technologies) and rhodamine phalloidin (Life Technologies) in PBS containing 10% normal goat serum at 4°C for 1 h in the dark. After five washes with PBS, the cover slips were mounted on slide glass with SlowFade Gold antifade reagent (Life Technologies). Confocal microscopy and differential interference contrast microscopy were conducted with a confocal laser scanning microscope (FV1000-D; Olympus Co., Tokyo, Japan).
To examine the effects of cytochalasin D, we differentiated Caco-2 in Biocoat cell culture inserts as mentioned above, inoculated the spores of K. septempunctata to Caco-2 cells in the presence of cytochalasin D at the concentration indicated, and measured the TER. Since cytochalasin D and DMSO showed weak toxicity on Caco-2 cells, TER was presented as a ratio to the culture without inoculation of spores.
Results and Discussion
Effects of K. septempunctata on the permeability across the Caco-2 cell monolayer
Caco-2 cell line has been widely used as an in vitro model for evaluating intestinal permeability because of its structural and functional similarity to mature intestinal epithelium (Lytton et al., 2005; Dickman et al., 2000). The cell monolayer permeability assay was performed to investigate the toxicity of K. septempunctata towards Caco-2 cells. We obtained P. olivaceus infected with K. septempunctata and purified spores from the trunk muscles. The spores were inoculated in Caco-2 cells, and the TER across the Caco-2 cell monolayer was measured as an indicator of permeability. The inoculation of purified K. septempunctata spores rapidly decreased TER by 80% within 1 h, indicating an increase in the permeability of the Caco-2 cell monolayer (Fig. 1). Our previous study showed that freezing inactivates K. septempunctata (Kawai et al., 2012); indeed, K. septempunctata purified from frozen P. olivaceus did not affect TER (Fig. 1). To determine whether K. septempunctata produces a toxic substance, spores were incubated in EDM at 37°C for 18 h, and the supernatant was recovered. When the culture medium from the Caco-2 cells was replaced with the supernatant from the spore, TER was not changed (Fig. 1). The inoculation of sonicated K. septempunctata spores also did not affect TER. These results suggested that K. septempunctata reduced TER by mechanisms other than production of toxin.

Increased permeability across the Caco-2 cell monolayer induced by Kudoa septempunctata spores. Differentiated Caco-2 cells were treated with purified live spores, frozen spores, sonicated spores, or culture supernatant from spores. Transepithelial electrical resistance (TER) was measured for up to 5 h. TER at time 0 is represented as 100%. Values are the mean±SD from three independent experiments.
Release of sporoplasms from spores
To further investigate whether K. septempunctata spores directly affect Caco-2 cells, we inoculated Caco-2 cells with spores and observed the cells under the microscope (Fig. 2). Within 1 h after inoculation, we observed the release of protoplasm from the spores on the surface of Caco-2 cells (Fig. 2, left panel), and this protoplasm, as well as the spores themselves, reacted with anti–Kudoa antiserum (Fig. 2, right panel). As it has been reported previously that some myxosporean parasites release sporoplasm into the host intestine (El Matbouli et al., 1998), we believe that this material is likely the sporoplasms of K. septempunctata.
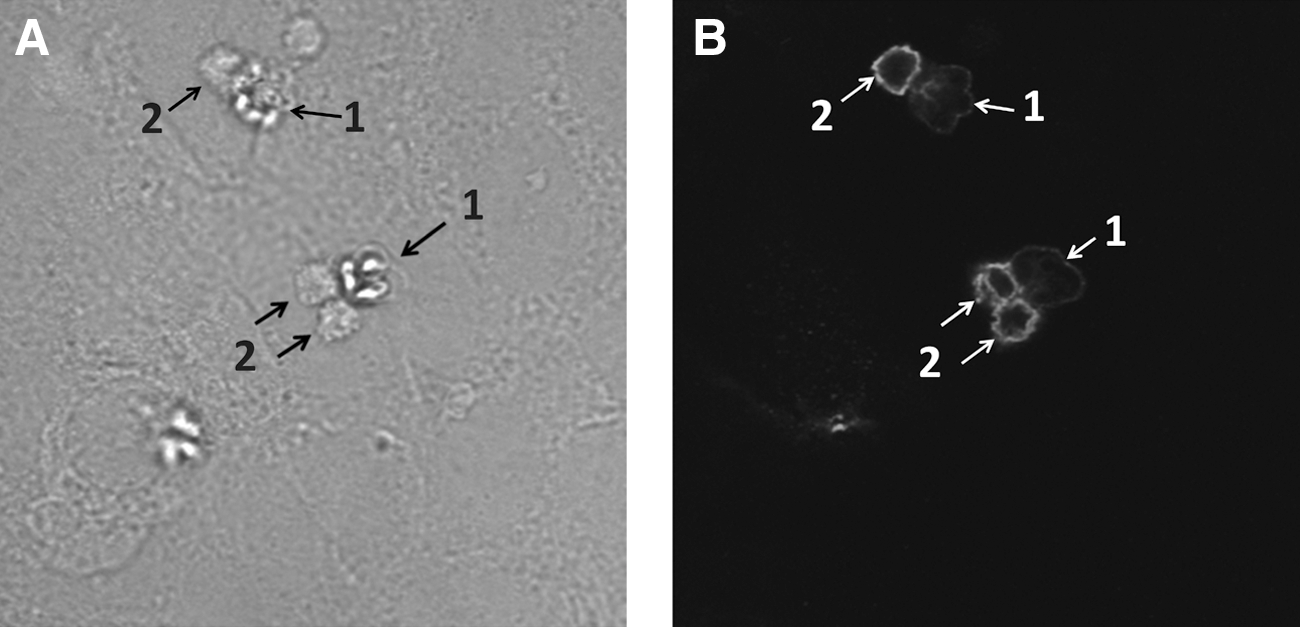
Release of sporoplasms from spores induced by inoculation of spores into Caco-2 cells. Differentiated Caco-2 cells were inoculated with Kudoa septempunctata spores and incubated for 1 h at 37°C. Samples were stained with α–K. septempunctata antiserum and rhodamine phalloidin, and analyzed by differential interference contrast microscopy
Next, we aimed to determine the stimulant responsible for the release of sporoplasms. Since the release of sporoplasm has been observed on Caco-2 cells cultured in the presence of FCS (T. Ohnishi, unpublished data), we first treated K. septempunctata spores with 10% FCS in PBS (Table 1). This treatment induced the release of sporoplasm, and the release was inhibited by the addition of a protease inhibitor cocktail (Table 1). These results indicated that one of the inducers is a protease present in FCS. We were not able to identify the specific protease in FCS, but a protease inhibitor cocktail blocking serine-, cysteine-, and trypsin-like protease activities inhibited sporoplasm release in FCS. Since one of the major proteases secreted in the intestine is trypsin, we treated K. septempunctata spores with trypsin and found that trypsin also stimulated sporoplasm release (Table 1). Interestingly, treatment with the protease pepsin did not induce the release of sporoplasm. These results suggest that some proteases secreted in the intestine can induce the release of sporoplasms from the spore. In our Caco-2 cell model, the cells were differentiated under serum-free conditions prior to experiments. However, it has been shown that the active forms of glycosylphosphatidylinisotol (GPI)–anchored trypsin-like proteases are expressed on the apical surface of Caco-2 cells (Friis et al., 2011). Caco-2 cells also express other proteases on their apical surface (Buzza et al., 2010; Wang et al., 2009). Therefore, we hypothesize that K. septempunctata spores utilize the proteases expressed on the apical surface of the intestinal cells to induce the release of sporoplasms in our Caco-2 cell model.
We used electron microscopy to investigate the role of sporoplasms in the pathogenesis (Fig. 3). The electron microscopy showed that K. septempunctata spores have an asymmetrically stellate shape in apical view (Fig. 3A) (Matsukane et al., 2010). When K. septempunctata spores were inoculated in Caco-2 cells, TEM confirmed the release of sporoplasms on the Caco-2 cell monolayer (Fig. 3B). Cross section analysis of the spore revealed two spaces for the sporoplasms (Fig. 3B). After release from the spore, the sporoplasm swelled and obtained an amoeba-like shape (Fig. 3C,D). Dense cytoskeletons were developed in the sporoplasm (Fig. 3E), and the sporoplasm developed pseudopodia (Fig. 3C). The discharge of polar filaments from the spore was also observed, and the polar filaments adhered to the cell surface (Fig. 3F).
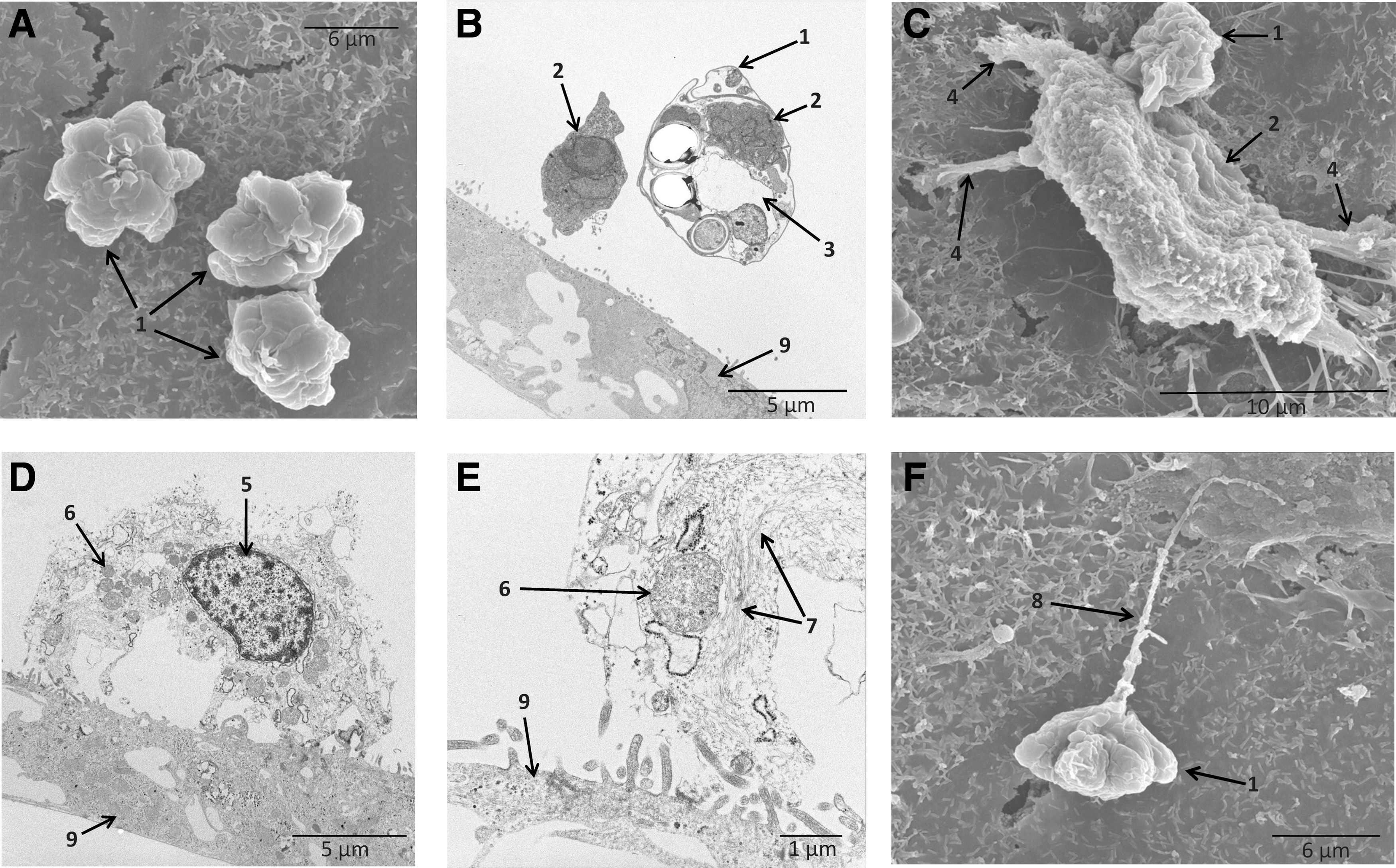
Morphological observations of Kudoa septempunctata by electron microscopy. Differentiated Caco-2 cells were inoculated with K. septempunctata spores and incubated for 1 h at 37°C. Samples were observed using both scanning electron microscopy (SEM) and transmission electron microscopy (TEM).
Sporoplasm invasion of Caco-2 cells
During infection, examination of cross section using TEM indicated that the sporoplasm intruded directly into the cytoplasm of Caco-2 cells (Fig. 4A). Examination of infecting sporoplasms by SEM suggested that this intrusion resulted in large holes on the Caco-2 cell surface (Fig. 4B). Following release from the spore, we observed that the cytoskeleton within sporoplasms became more defined (Fig. 4C). This reflects the condensation of actin filaments (Alama-Bermejo et al., 2012), likely to aid in the movement of the sporoplasm in the Caco-2 cell. We hypothesized that inhibiting actin polymerization in the sporoplasm would prevent the movement of sporoplasm, resulting in the inability to release from spores or invade cells. When we inoculated K. septempunctata in Caco-2 cells in the presence of cytochalasin D, treatment with 2 μM cytochalasin D completely inhibited the release of sporoplasm from spores (Fig. 5A, lower panel). In contrast, 1 h after infection without cytochalasin D, the sporoplasms had reached the basolateral side of the Caco-2 cells, as evidenced by confocal microscopy (Fig. 5A, upper panel). The reduction of TER induced by K. septempunctata was suppressed by cytochalasin D in a dose-dependent manner and was completely inhibited at a concentration of 2 μM (Fig. 5B). Collectively, our results demonstrate that invasion by K. septempunctata sporoplasms eliminated TER across the Caco-2 cell monolayer. Stopping cell invasion either by inactivating the spores or by inhibiting the release of sporoplasm prevented loss in TER. The sporoplasm of K. septempunctata very quickly intruded into Caco-2 cells and reached the basolateral side of Caco-2 cells within 1 h of infection. This rapid invasion coincides with the quick reduction of TER and the short lag phase before the appearance of clinical symptoms.

Cell damage caused by sporoplasm invasion of Caco-2 cells. Differentiated Caco-2 cells were inoculated with Kudoa septempunctata spores and incubated for 1 h at 37°C. Samples were observed using both scanning electron microscopy (SEM) and transmission electron microscopy (TEM).

Effect of increasing concentrations of cytochalasin D on sporoplasm-mediated reduction in the transepithelial electrical resistance.
Conclusion
Our results demonstrate that the invasion of human epithelial cells by K. septempunctata sporoplasms occurs quite rapidly, resulting in the termination of monolayer confluence as illustrated by the rapid loss of the TER due to severe damage of individual cells within the monolayer. Rapid invasion of gut epithelial cells by K. septempunctata sporoplasms may be a contributing factor to the diarrhea associated with this pathogen, although additional work will be required to further elucidate the mechanisms of pathogenesis in K. septempunctata.
Footnotes
Acknowledgments
We thank Yutaka Fukuda, Oita Prefectural Agriculture, Forestry and Fisheries Research, and Mitsuru Ototake and Hiroshi Sako, National Research Institute of Aquaculture, for valuable advices. This research was supported by the Ministry of Health, Labour and Welfare, Japan (Health Labour Sciences Research Grant H23-shyokuhin-ippan-007).
Disclosure Statement
No competing financial interests exist.