
Abstract
Campylobacters have developed a number of mechanisms for responding to environmental conditions, although the different virulence properties of these cells following exposure to stress are still poorly understood. We analyzed in vitro stress responses and the consequent in vivo modulation of Campylobacter jejuni pathogenicity in BALB/c mice, as a result of the exposure of the C. jejuni to environmental stress (starvation, oxidative stress, heat shock). In vitro, the influence of starvation and oxidative stress was milder than that of heat shock, although the majority of the stress conditions influenced the survival of C. jejuni. During starvation, C. jejuni viability was maintained longer than its culturability. Additionally, starvation elicited transformation of stressed bacteria to coccoid forms. In contrast, bacteria exposed to oxygen remained culturable, but their viability decreased. Pre-starvation did not contribute to improved survival of C. jejuni cells during oxygen exposure. Changes in bacteria numbers and the levels of several cytokines (interleukins 6 and 10, tumor necrosis factor-α, interferon-γ) were followed in vivo, in liver homogenates from the mice intravenously infected with either control (untreated) or stressed C. jejuni. The systemic infection with the control or stressed C. jejuni occurred with different production dynamics of the cytokines investigated. Starvation was the most powerful stress factor, which significantly decreased infectious potential of C. jejuni during the first 3 days postinfection. The most pronounced differences in cytokine production were found in interferon-γ and interleukin-10 production, which indicates that these have roles in the immune response to C. jejuni infection. These in vivo studies of environmental impact on bacterial virulence reveal that microbial adaptation during stress challenge is crucial not just for pathogen survival out of the host, but also during host–pathogen interactions, and thus for the bacterial pathogenicity.
Introduction
H
Animal models are indispensable for information about bacterial virulence properties in vivo. Numerous in vitro experiments have demonstrated that C. jejuni can invade intestinal epithelial cells and can also survive for long periods inside both phagocytic and epithelial cells (Chang and Miller, 2006; Rubeša Mihaljević et al., 2007; Hermans et al., 2011). Infection of mice followed by systemic C. jejuni spread and the infection of internal organs has indicated that the mouse is a useful in vivo model to demonstrate bacterial dissemination and tissue invasion (Vučković et al., 1998).
A significant part of C. jejuni pathogenesis is connected with the effects of lipopolysaccharides, which are responsible for the production of several cytokines (Hu et al., 2006). These polypeptides have important roles in the pathogenesis of Gram-negative infections, when both pro-inflammatory and anti-inflammatory cytokines are produced (Dinarello, 1996; Rukavina et al., 2006). Pro-inflammatory cytokines, such as interleukin (IL)-1β, tumor necrosis factor (TNF)-α, and IL-6, participate in the induction of acute-phase reactions, including fever. A number of feedback mechanisms have synergistic or antagonistic influences on the production and activity of individual members of the cytokine network (Jagusztyn-Krynicka et al., 2009; Zídek et al., 2009). Verhoeff-Bakkenes et al. (2008) showed a correlation between culturability properties and induction of IL-8 secretion using the INT-407 human intestinal epithelium cell line. However, it is still not known whether in vivo host immune responses, such as cytokine production, are affected after infection with bacteria that have been exposed to environmental stress.
We conducted a study to investigate whether in vivo modulation of C. jejuni virulence is altered in response to selected stress conditions. First, we studied bacterial stress responses in terms of physiological (culturability, viability) and morphological changes. To better understand the involvement and interactions of the different defense mechanisms on bacterial survival and infectivity, we also monitored the bacterial burden and local cytokine production in the liver of BALB/c mice following their infection with the stressed C. jejuni.
Materials and Methods
Bacterial growth and stress conditions
C. jejuni K49/4 were isolated from poultry meat in our laboratory and identified phenotypically and by multiplex polymerase chain reaction (Zorman and Smole Možina, 2002). All of the experiments used exponential phase cultures of the isolate following microaerobic growth (5% O2, 10% CO2, 85% N2) in Preston broth (Oxoid, Hampshire, UK) at 42°C for 9 h (Klančnik et al., 2006, 2009). For the stringent response, the C. jejuni cells were harvested by centrifugation (12,000×g for 5 min at 4°C), washed, resuspended in Ringer's solution supplemented with 5 mM KH2PO4 (Kemika, Zagreb, Croatia), incubated microaerobically for 5–72 h at 42°C, and then exposed to starvation. For pre-starvation, the C. jejuni cells were prepared as described previously and incubated for 5 h at 42°C before the subsequent oxidative stress. To produce the oxidative stress, these C. jejuni cells were exposed to oxygen at atmospheric concentration for 5–72 h. To produce the heat-shock stress, the temperature was shifted to 55°C for 3 min, and the cells were then cooled on ice before analysis. Untreated C. jejuni from exponential growth-phase cultures were used as the simultaneous controls for each of the stress conditions analyzed.
Culturability and viability assay
Culturability was determined as the colony-forming units per milliliter (CFU/mL), and viability was determined using the LIVE/DEAD BacLight system (L-7012; Molecular Probes, Eugene, OR) under an Eclipse TE300 microscope (Nikon, Tokyo, Japan), as previously described (Klančnik et al., 2006). Viability is given as the percentages of viable cells in relation to the total number of cells obtained, as determined under the microscope for 20 randomly chosen visual fields per filter. The numbers of coccoid and spiral cells were also determined. Of note, although it has been suggested that there are limitations to the use of microscopy for the determination of viable coccoid cells, which might thus affect the interpretation of the results obtained, we have previously demonstrated that these can be overcome. This was achieved by using a well-known commercial system for the visualization of the live bacteria. Also, we have confirmed the quality of this test previously in numerous systems that have compared the results obtained with flow cytometry (Rubeša Mihaljević et al., 2005; Klančnik, 2006). All of the experiments were independently repeated three times. C. jejuni morphology was assessed using transmission electron microscopy (Philips CM 100, Philips Electronics N.V. Eindhoven, The Netherlands). The cells were prepared as reported previously (Klančnik et al., 2009).
In vivo experiments
Male BALB/c (H-2d) mice 8–12 weeks of age were used in all of the in vivo experiments. These were obtained from the Central Animal Facility of the Medical Faculty, University of Rijeka, Croatia, and were given standard laboratory rodent food (Mucedola, Milan, Italy) and water ad libitum. The experiments were conducted in accordance with the guidelines of the International Guiding Principles for Biomedical Research Involving Animals (NCR, 2004). The Ethical Committee at the University of Rijeka approved all of the animal experiments described here. The mice were infected intravenously via the lateral tail vein with a single dose (200 μL) of 0.5–1.0×109 CFU of untreated (control) or stressed C. jejuni cells, as determined by the turbidity of the bacterial suspension and confirmed retrospectively by plating the inoculum on blood agar and microaerobic incubation for 48 h at 42°C. At 1, 3, and 8 days postinfection, the mouse livers were removed aseptically and dissected from the surrounding tissue, and the C. jejuni CFU in the livers were determined as previously described (Vučković et al., 1998). For the organ C. jejuni burden experiments and cytokine analyses, two sets of mice were infected, with at least three per group. The C. jejuni burden experiments were repeated three times and the data from all of the replicate experiments were pooled and are presented as means±standard deviations.
Cytokine levels
For the determination of the levels of the cytokines, the livers were removed aseptically, dissected from the surrounding tissue, and collected in preweighed sterile vials (Eppendorf AG, Hamburg, Germany) on ice. The weights of the livers were recorded on an electronic balance (PB602-S; Mettler, Switzerland). The livers were then frozen in liquid nitrogen and stored at −80°C. Later, these frozen livers were thawed on ice and homogenized in ice-cold phosphate-buffered saline (5 mL per 1 g tissue, wet weight) with a hand-held tissue homogenizer. They were then centrifuged (14,000×g for 10 min at 4°C) to precipitate the cell debris as a pellet. The supernatants were stored at −20°C until they were assayed for the cytokine levels.
The levels of IL-6, IL-10, TNF-α, and interferon (IFN)-γ were determined using mouse cytokine enzyme-linked immunosorbent assay kits (Thermo Scientific, Pierce Products, Rockford, USA). The assays were performed in duplicate according to the manufacturer instructions, and the results are expressed as pg/mL tissue homogenate. The sensitivity levels were: IL-6, 7 pg/mL; IL-10, 12 pg/mL; TNF-α, 9 pg/mL; and IFN-γ, 10 pg/mL. Control mice of the same age and sex were injected with sterile saline. All of the experiments were independently repeated three times, and the data are presented as means±standard deviations.
Statistical analysis
The differences in the bacterial counts and the cytokine levels between all of the conditions of the experimental groups were calculated using the Kruskal–Wallis test, while the Mann–Whitney test was used to define the differences between all of the pairs of groups. All statistical values were considered significant at a p level of 0.05. Statistical analysis was performed using SPSS 15.0 for Windows (Statsoft Inc., Tulsa, OK).
Results
Campylobacter survival and morphology under stress conditions
Figure 1A shows that although the total viability of the C. jejuni decreased with incubation time, the percentage of coccoid cells increased at the same time. C. jejuni starvation strongly influenced their culturability, while their viability remained stable until the end of experimental period (72 h), when there were no culturable C. jejuni and there was a marked decline in viable cells (Fig. 1B). Of note, potential limitations of microscopy for the determination of viable coccoid cells were avoided by using a well-known, previously validated commercial system for the visualization of the live bacteria (see Materials and Methods). Morphological changes were already seen after 5 h of starvation (Fig. 2), when the number of coccoid cells began to increase (Fig. 1B). On the contrary, after exposure to oxygen, the C. jejuni viability was more affected than their culturability (Fig. 1C). Here, the morphological changes were mild, although they were seen continuously throughout the exposure period, with the spiral, shorter spiral, and coccoid forms evident (Fig. 2). C. jejuni cells that were pre-starved for 5 h were even more susceptible to this oxygen stress. As seen in Figure 1D, here the number of viable or culturable cells under oxygen exposure was already reduced to below the detectable level after 24 h. The bacterial morphology was also more affected in these pre-starved C. jejuni cells under oxygen stress (Fig. 2). The temperature shift to 55°C for 3 min resulted in an immediate reduction in the C. jejuni culturability and viability (Fig. 1E), which were accompanied by prominent morphological changes (Fig. 2).

Culturability (line) and viability (bars) of control Campylobacter jejuni
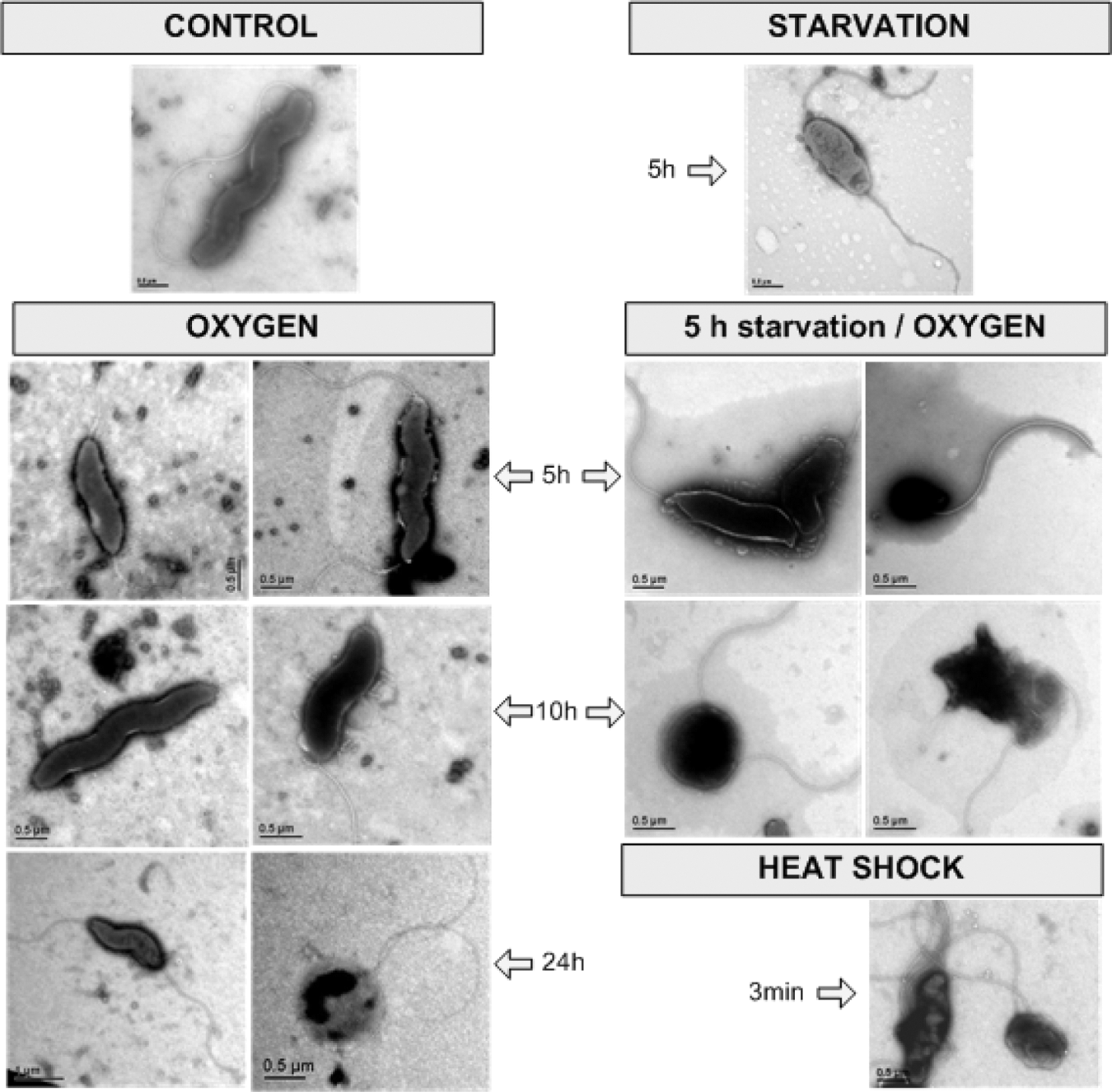
Electron microscopy showing control Campylobacter jejuni and the effects of starvation, oxygen exposure (with and without pre-starvation), and heat shock on bacterial morphology. Bars: as indicated.
Modulation of Campylobacter virulence under stress conditions
To further define the influence of sublethal stress (5 h starvation, 5 h oxygen) on C. jejuni virulence, we followed the course of C. jejuni infection in an animal model. The heat-shocked C. jejuni cells were not used in these in vivo experiments because of their reductions in culturability and viability and marked changes in morphology that were caused within 3 min of the heat exposure. The nontreated and starvation- and oxygen-exposed C. jejuni efficiently reached the mouse livers after the intravenous injections (Fig. 3). Here, starvation was the more powerful of these two stress factors for reduction in the infectious potential of these C. jejuni. As shown in Figure 3, the liver colony counts on the first and third days postinfection were significantly lower in the mice infected with the starved C. jejuni, in comparison to the control and oxygen-stressed C. jejuni. No differences were seen in the numbers of C. jejuni cells isolated from the livers of mice infected with the untreated and oxygen-exposed campylobacters.
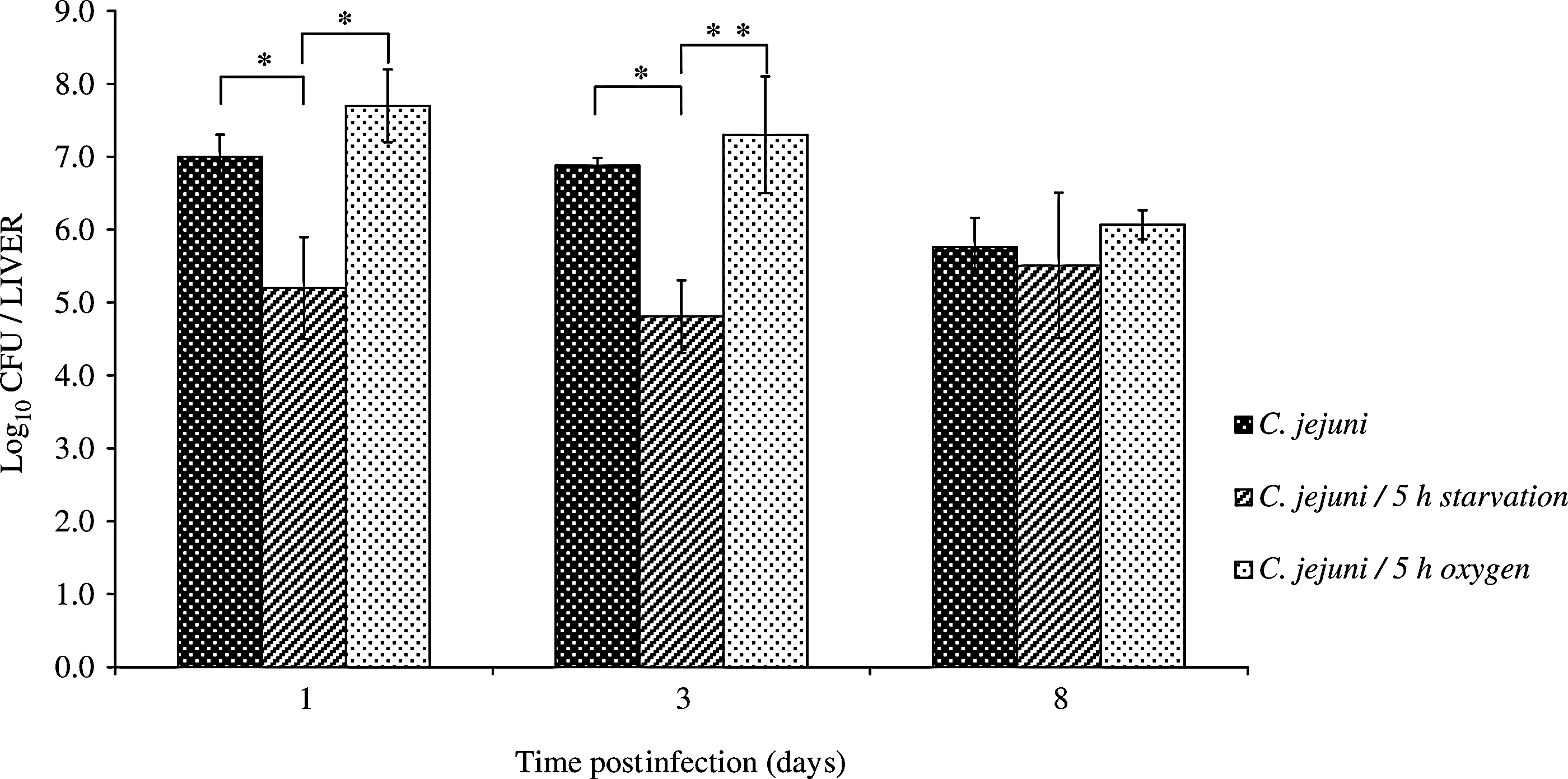
The bacterial counts in the liver of BALB/c mice intravenously infected with control or stressed Campylobacter jejuni. Data are presented as log10 of mean values±standard deviations of C. jejuni colony-forming units (CFU)/liver (*p≤0.05; **p=0.054).
The levels of the different cytokines in the liver homogenates were analyzed at selected times, as the responses to the infections caused by the untreated and stress-exposed C. jejuni. The total liver IL-6 levels showed similar patterns in all of the experimental groups. The highest IL-6 levels were seen on day 1 postinfection, although these did not reach significance over the control (uninfected) mice; these levels then decreased thereafter. By day 8, the IL-6 levels in the control mice (infected with untreated C. jejuni) had returned to baseline, while they were significantly elevated in the animals treated with the stressed C. jejuni. Thus, significant differences in the IL-6 levels were noted in the groups infected with 5-h-starved and 5-h-oxygen-exposed C. jejuni, in comparison to the baseline values (Fig. 4A).
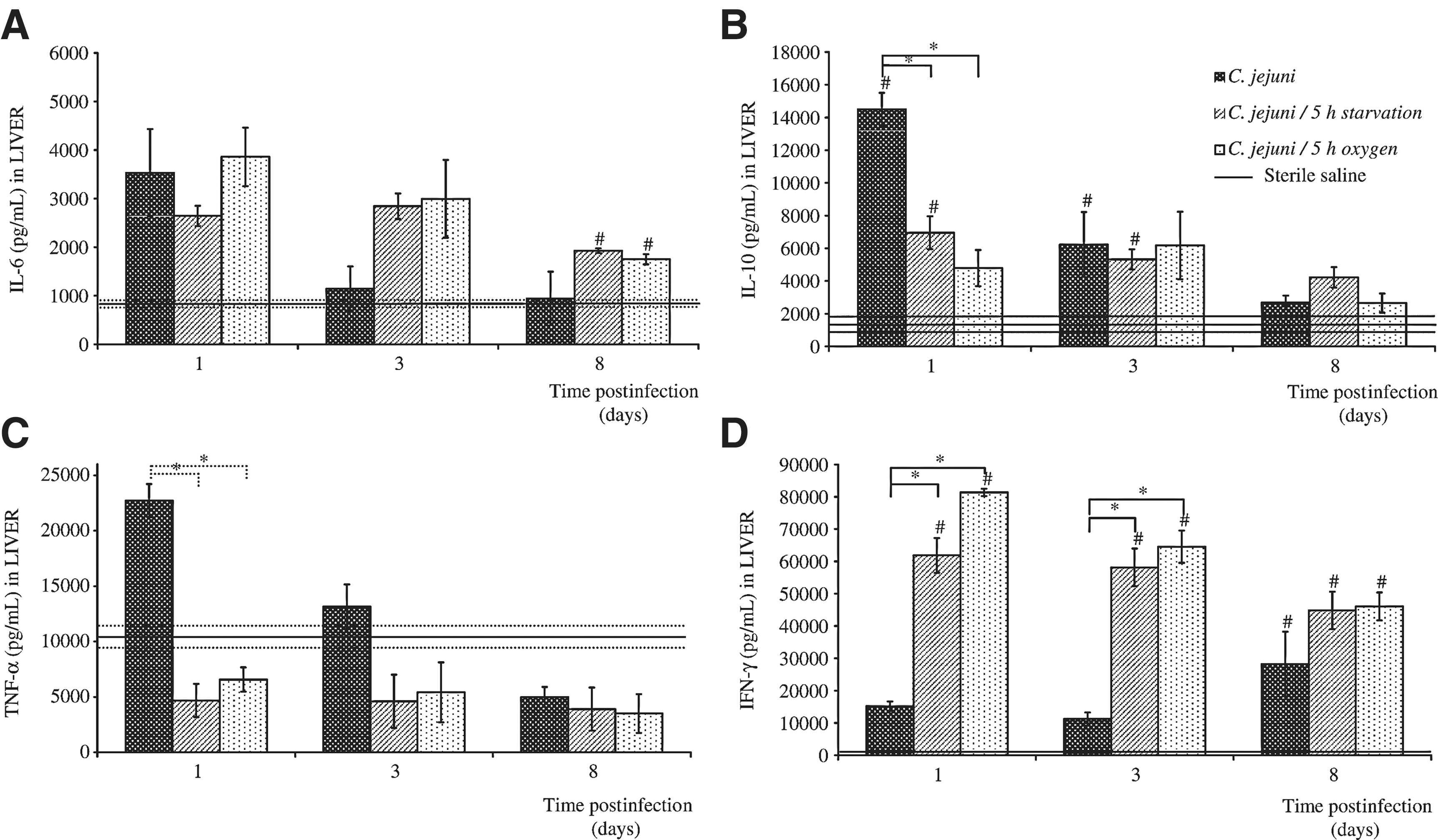
The levels of interleukin (IL)-6
In the mice infected with the untreated C. jejuni, IL-10 also reached greatly increased maximal levels in the liver homogenates on the first day postinfection, which were significantly higher than for saline-treated mice (by ∼10-fold) and for the mice infected with either of the stressed C. jejuni. These liver IL-10 levels decreased thereafter. On day 3, the IL-10 levels were still significantly higher in the livers of the mice infected with control or starved C. jejuni, in comparison to the basal value (Fig. 4B).
In the case of TNF-α, when compared to the starved and oxygen-stressed C. jejuni, significantly higher levels were detected on the first day postinfection in the liver homogenates of the mice infected with the untreated C. jejuni. However, due to the variability of the TNF-α data at this time, there was no significant difference between the TNF-α production in the liver homogenates of the mice infected with the treated C. jejuni and the mock-infected controls (Fig. 4C).
The liver homogenate IFN-γ levels in the mice infected with the untreated C. jejuni were not significantly different from the basal levels until day 8 postinfection. However, IFN-γ showed the greatest induction for all of these cytokines following infection with the C. jejuni exposed to stress, either as induced by starvation or oxygen, with the IFN-γ levels remaining significantly higher than the baseline throughout the whole experimental period. Thus, the mice infected with these stressed C. jejuni had significantly higher IFN-γ levels in their livers than the mice infected with the untreated control C. jejuni (Fig. 4D).
Discussion
Food processing, distribution, and storage rely on well-placed deleterious stress conditions, as hurdles, that either inhibit or inactivate microbial contaminants in food systems. However, many foodborne bacteria have evolved sophisticated strategies and mechanisms to deal with these environmental stress conditions, and have consequently become more resistant to these stresses, and potentially cross-protected to other stresses. Thus, microbial adaptation to harsh environmental conditions that can impact bacterial survival might compromise food safety, and such enhanced bacterial pathogenicity can lead to increased incidence of disease. Research into food safety is therefore dependent on our knowledge of the specific foodborne pathogens and their pathogenesis in the susceptible host.
Like other foodborne bacteria, C. jejuni is exposed to various stress conditions in food-processing environments, as well as in their host (Atack and Kelly, 2009). We have analyzed here the C. jejuni stress responses after their exposure to selected environmental stress, following their survival and their morphological changes, as well as the subsequent establishing of systemic mouse infections and the stimulation of liver cytokine production.
In this study, starvation had effects on the C. jejuni culturability, rather than their viability, which corresponds to a continuous increase in the number of coccoid cells. The rapid decline in culturability and the slower decline in viability suggest that these starved C. jejuni cells went into their state. As is known, the conversion to the viable but nonculturable form and the transition to a coccoid form are two different processes, although they can also be related (McDougald et al., 1998; Klančnik et al., 2009). In contrast, when the C. jejuni cells were tested for their survival under oxidative stress conditions, their culturability was essentially not affected. However, prolonged oxygen exposure led to decreased C. jejuni cell numbers, which were reduced to undetectable levels.
Considering the known importance of sublethal prestress starvation for improved heat resistance (Klančnik et al., 2006), we aimed to estimate the influence of 5 h starvation on the survival of C. jejuni under subsequent oxidative stress. Contrary to improving heat resistance, sublethal cell starvation did not enhance the survival of C. jejuni during the following oxygen exposure. In fact, our data show that oxygen exposure is a strong stress for prestarved cultures.
Our data confirm the high susceptibility of C. jejuni to heat shock. Changes in C. jejuni cell morphology were seen immediately after this heat exposure, which were accompanied by decreased culturability and viability.
In our previous studies, we reported that sublethal environmental stress factors, like 5-h starvation and 5-h oxygen exposure, might have implications on the pathogenesis-related functions of C. jejuni, such as their adhesion, invasion, and intracellular survival (Rubeša Mihaljević et al., 2007; Klančnik et al., 2009; Šikić Pogačar et al., 2009, 2010). Knowing that adhesion and invasion of pathogenic bacteria are the important initial steps of bacterial infection, in the present study we focused on a mouse model of campylobacteriosis. We examined the influence of these same stress conditions on C. jejuni pathogenicity, following the systemic infections cause by these stressed bacteria in vivo in the experimental mice, as well as the mouse host response according to the analysis of liver cytokine levels.
After intravenous inoculation, regardless of being stressed or not, all campylobacters established liver infections. Starvation significantly decreased the infectious potential of these C. jejuni, resulting in the lowest numbers of C. jejuni in the liver. This can be explained by the probable transformation of these cells into the dormant form, or the viable but nonculturable form, as indicated by the decline in culturability and the altered cell morphology. Using in vitro cell culture models, we previously reported that 5-h starvation has a strong impact on C. jejuni adhesion and invasion in enteric (Caco-2) cells (Rubeša Mihaljević et al., 2007), as well as in macrophages (J774) (Šikić Pogačar et al., 2009), and thus this might affect the establishment of infection in vivo. In contrast to the strong influence of starvation, oxidative stress neither affected the C. jejuni infectivity nor reduced the C. jejuni load in the mouse liver, in comparison to the untreated C. jejuni. This is in agreement with the moderate changes in cell morphology and the preserved viability of the C. jejuni exposed to 5 h of oxygen. This finding is also supported by our previous in vitro studies (Rubeša Mihaljević et al., 2007; Šikić Pogačar et al., 2009, 2010), which showed that 5-h exposure to oxygen enhances the binding and invasion properties of C. jejuni. However, in the present study, on day 8 postinfection, there were no significant differences of the C. jejuni growth in the liver among these three groups.
During infections, cytokines have central roles in the regulation of immune and inflammatory responses, as well as in many other host functions. Cytokines, which can be pro-inflammatory or anti-inflammatory, are the main regulators of local tissue immune responses. This prompted us to determine the cytokine responses in the liver of these mice infected with untreated and stress-exposed C. jejuni. The pro-inflammatory cytokines, such as IL-6, TNF-α, and IFN-γ, are usually produced in the organs, including the liver, under nonlethal and lethal infections with different pathogens. IL-6 is a multifunctional cytokine that has been described as having both pro-inflammatory and anti-inflammatory effects, as well as being involved in a variety of immune responses. Here, there were increased levels of IL-6 in the mouse livers, although there were no significant differences among the groups. TNF-α is a potent pro-inflammatory cytokine that is released primarily from stimulated macrophages and that is known to induce the production of IL-6. However, in our model, all of the TNF-α levels were below baseline, apart from an increase in the TNF-α levels in the mouse livers on the first day postinfection with untreated C. jejuni, which was not significantly different from the control (uninfected) mice. The production of TNF-α is regulated by the anti-inflammatory effects of IL-10 (de Waal Malefyt et al., 1991). A lack of TNF-α can be explained by high levels of IL-10. As only live bacteria, and not dead bacteria, can induce IL-10 production, this might explain the higher levels of IL-10 in the livers of the mice infected with the untreated C. jejuni. It is possible that the high IL-10 levels in the mouse livers in response to the live and viable untreated C. jejuni represent a consequence of the immune system trying to counteract and suppress the increased inflammation, and to antagonize IFN-γ production. It follows that the lower liver IL-10 levels in the mice infected with C. jejuni after their exposure to starvation or oxygen leads to clear increases in the levels of IFN-γ. C. jejuni can induce both pro-inflammatory and anti-inflammatory cytokines. IFN-γ and IL-10 appear to have the main roles here. The IL-10 might have contributed to the resolution of the inflammation and inhibition of the production of the pro-inflammatory IFN-γ in the mice infected with the untreated C. jejuni. However, it appears that stressed bacteria can induce higher IFN-γ production, at least over the first few days postinfection. Whether this affects the severity of the disease or the histopathological changes in the liver requires further study.
Conclusions
Our findings on the in vitro and in vivo modulation of the C. jejuni viability and pathogenicity characteristics in response to environmental stress factors provide important insights for a better understanding of the C. jejuni infective cycle. They also thus contribute further toward the methods relating to improved safety in the food production/supply chain.
Footnotes
Acknowledgments
This study was supported by the Ministry of Higher Education, Science and Technology of the Republic of Slovenia, by the Z1-2190 post-doctoral project of A.K., by a bilateral Slovenian-Croatian project, and through research projects financed by the Croatian Ministry of Science, Education and Sports (062-0621273-1235, 062-0621273-0949).
Disclosure Statement
No competing financial interests exist.