
Abstract
An epidemiological survey on human norovirus (NoV)–associated gastroenteritis was conducted to clarify the prevalence of NoV infections in children and adults in Korea. Recombinant capsid proteins from three major NoV genotypes (GI-4, GII-3, and GII-4) were expressed using a baculovirus expression system, and the morphology and antigenicity of self-assembled virus-like particles were then confirmed by electron microscopy and Western blotting with a NoV-specific antibody. To determine seroprevalence, an enzyme-linked immunosorbent assay was performed to detect antibodies against virus-like particles antigen in 346 serum specimens collected from persons who visited five public heath care centers for regular physical examination in Jeollanam-do, Korea, between 2005 and 2006. The seroprevalence of immunoglobulin G antibodies against the GI-4, GII-3, and GII-4 NoV genotypes was 84.1%, 76.3%, and 94.5%, respectively. A rapid decrease in seroprevalence occurred after birth, with the lowest levels observed in the <23-month age group, and a steep increase in seroprevalence occurred in early childhood, reaching 60.5% for GI-4, 65.1% for GII-3, and 90.7% for GII-4 at age 2–5 years, and over 80% for all three genotypes in subjects aged 20 years or older. The seroprevalence of different NoV genotypes statistically differed across the age groups (p<0.01).
Introduction
NoVs are small, round viruses 27–35 nm in diameter with surface structures that are distinct from morphologically typical caliciviruses when observed with an electron microscope (Kapikian et al., 1972). The NoV genus is divided into five genogroups (GI to GV) on the basis of genetic diversity, and two genogroups (GI and GII) are major pathogens in humans (Kageyama et al., 2004; Zheng et al., 2006). Seroepidemiological studies of NoV can be informative regarding the prevalence of infections in the population, but have been restricted by the absence of a viral cultivation system for NoV. Therefore, several studies have investigated the seroprevalence of NoV by using virus-like particles (VLPs) formed from self-assembled antigens expressed in a baculovirus expression system because of their morphological and antigenic similarities to native NoV.
In the present study, recombinant capsid proteins from three NoV strains that are highly prevalent in Korea were expressed using a baculovirus expression system, and the seroprevalence of antibodies against these NoV strains were determined by enzyme-linked immunosorbent assay (ELISA) with the VLP antigens. The present study is the first to investigate the prevalence of infections with various NoV genogroups in Korea, with the aim of providing basic epidemiological information that will aid in the understanding of NoV infections in the Korean population and worldwide.
Materials and Methods
Molecular cloning and expression of the NoV capsid protein
Viral RNA was extracted from stool specimens containing NoV GI-4, NoV GII-3, and NoV GII-4 with the QiaAmp viral RNA kit (Qiagen). Oligo dT (Invitrogen) and Superscript II RNase H (Life Technologies) were used according to the manufacturer's instructions to obtain cDNA. The entire ORF2 gene, which encodes the capsid protein, was generated using the primer pairs shown in Table 1. The amplified product was sequenced, and inserted into the pENTR/D-TOPO vector (Invitrogen). For recombinant baculovirus preparation, the BacHTS System (Newgex, Korea) was used. In brief, the entry clone and vBacHTS-HisFlag viral DNA (having N-terminal six histidine and flag epitope fusion tag coding sequence “MHHHHHHGSDYKDDDDKGTTSLYKKAGSAAAPFT-”) were combined in vitro with LR clonase (Invitrogen). The reaction product was transfected into Sf21 cells with Lipofectin reagent (Invitrogen) to produce the recombinant baculovirus (Jiang et al., 1992).
Bases used for directional cloning are underlined.
Three recombinant NoV capsid antigens were purified from Sf21 (Clontech) insect cells infected with recombinant baculoviruses carrying the NoV ORF2 gene as described in previous investigations (Jiang et al., 1992; Jiang et al., 1995) with minor modification. In brief, 1.0×10^8 Sf21 cells were seeded into 850-cm2 roller bottles (Falcon) and attached 12 h, cultured old media was discarded, infected with 10 mL of recombinant baculovirus stock (1×10^8 plaque-forming unit/mL) for 2 h, 200 mL of fresh SF900II-SFM media containing 5% FBS (Invitrogen) was added and cultured 60–72 h on roller apparatus (Bellco) rotating 0.25 rpm in a 28°C incubator. Harvested cells from each roller bottle (200 mL culture) were resuspended in 6-mL PBS supplemented with complete proteinase inhibitor cocktail (Roche) and sonicated on ice for 30 s (0.5-s pulse 40 times). After centrifugation of 12,000 rpm for 15 min, supernatant was applied to 20–50% sucrose step gradient ultracentrifugation (Beckman, SW28 roter) at 25,000 rpm for 2 h. The morphology and antigenicity of recombinant NoV was confirmed by electron microscopy and Western blot with horseradish peroxidase–conjugated NoV-reactive polyclonal antibody (R-Biopharm, Germany).
Western blot
The purified capsid protein was resolved by a sodium dodecyl sulfate-polyacrylamide gel electrophoresis with 10% acrylamide gels and transferred to a nitrocellulose membrane by using the iBlot™ Gel Transfer Unit (Invitrogen, USA). After transfer of the proteins, the nitrocellulose membrane was washed in TTBS blocking solution (TBS [NaCl, 0.5 M; Tris-HCl, 20 mM; pH 7.5] plus Tween 20, 0.05% [vol/vol]) at 25°C. The capsid proteins were detected using the anti-NV antibody peroxidase conjugate in a commercial ELISA kit (NV-AD Seiken NoV antigen ELISA) (Denka-Seiken, Japan). Peroxidase-antibody conjugates were detected with diaminobenzidine.
Serum specimens
Blood specimens were collected from children and adults undergoing regular physical examination at five health care centers in Jeollanam-do, Korea, between 2005 and 2006. All samples were obtained with informed consent. A total of 346 serum samples were collected from individuals without gastrointestinal disease, ranging in age from birth to over 70 years. Sera were stored at −20°C before testing.
ELISAs to detect antibodies against NoV
A previously reported ELISA protocol was modified to detect antibodies against NoV (Jiang et al., 1992; Jiang et al., 1995). Briefly, each well of a 96-well maxisorp immunoplate (Nunc, Denmark) was coated with 100 μL of 5 μg/mL purified rNoV diluted in 0.01 M PBS, pH 7.4. The antigen-coated plate was incubated overnight at 4°C, washed once (200 μL per well) with PBS containing 0.05% Tween 20 (TW-PBS), and then blocked with 5% Blotto (Carnation nonfat milk) in PBS for 1 h at 37°C. After washing twice (200 μL per well) with TW-PBS, 100-μL samples of human serum diluted 1:2,000 in 0.05% TW-PBS were added to the wells, and the plate was incubated for 2 h at 25°C. After six washes with 0.05% TW-PBS, 100 μL of horseradish peroxidase–conjugated goat anti-human immunoglobulin G (Abcam, England) diluted 1:4000 in 0.05% TW-PBS was added to each well, and the plate was incubated for 2 h at 25°C. After a final six washes with TW-PBS, the bound antibody was detected by addition of 3,3,5,5-tetramethylbenzidine (TMB, GIBCO BRL, Gaithersburg, MD). After 10-min incubation at room temperature, a solution of 1.0 N H2SO4 was added to stop color development. An Emax microtiter plate reader (Molecular Devices, Sunnyvale, CA) was used to quantify the reactivity of each well at optical density at 450 nm. The optimal concentrations of the coating antigens and the secondary antibody used in the assays were determined by titration. A specimen with an OD450>0.2 was considered to be positive. This cutoff value of an ELISA is verified by a comparison with a group of negative control serum specimens.
Statistical analysis
Chi-square analysis was performed to compare the antibody detection rates between the three recombinant ELISAs in different age groups.
Results
Molecular cloning and expression of NoV capsid proteins
The ORF2 sequences of the three NoV strains were predicted to encode proteins of 544, 548, and 540 amino acids in length. Considerable diversity was observed across genotypes, with 40.9–43.7% homology at the deduced amino acid level. The two genotype 2 strains were 66.5% homologous at the deduced amino acid level (Table 2). Coomassie blue staining of 10% denaturing acrylamide gels indicated that Sf9 cells infected with recombinant baculovirus carrying the NoV ORF2 gene released recombinant capsid protein of approximately 60 kDa (excluding tagging proteins) into the supernatant. The antigenicity of expressed proteins was confirmed by Western blotting with NoV-specific antibodies, as shown in Fig. 1. With performance of electron microscopy, virus particles purified by sucrose gradient centrifugation self-assembled to form VLPs of 35–40 nm in size were identified (Fig. 2).
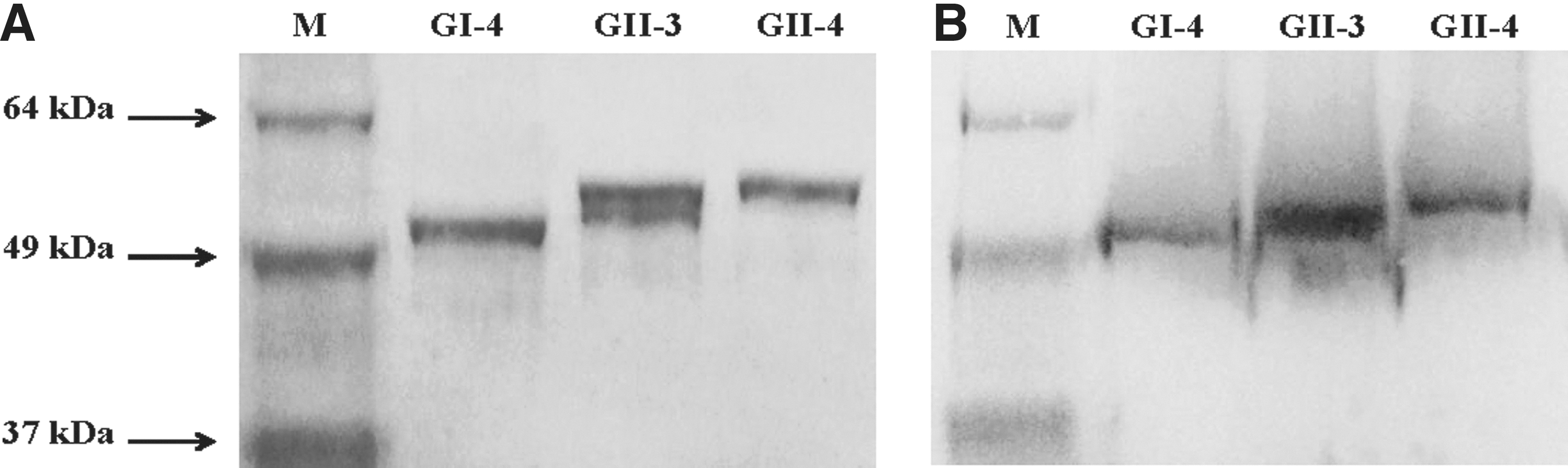
Recombinant virus-like particles (VLPs) of noroviruses (NoV) electrophoresed on a sodium dodecyl sulfate–polyacrylamide gel and immunoprecipitation with NoV-specific antibodies.
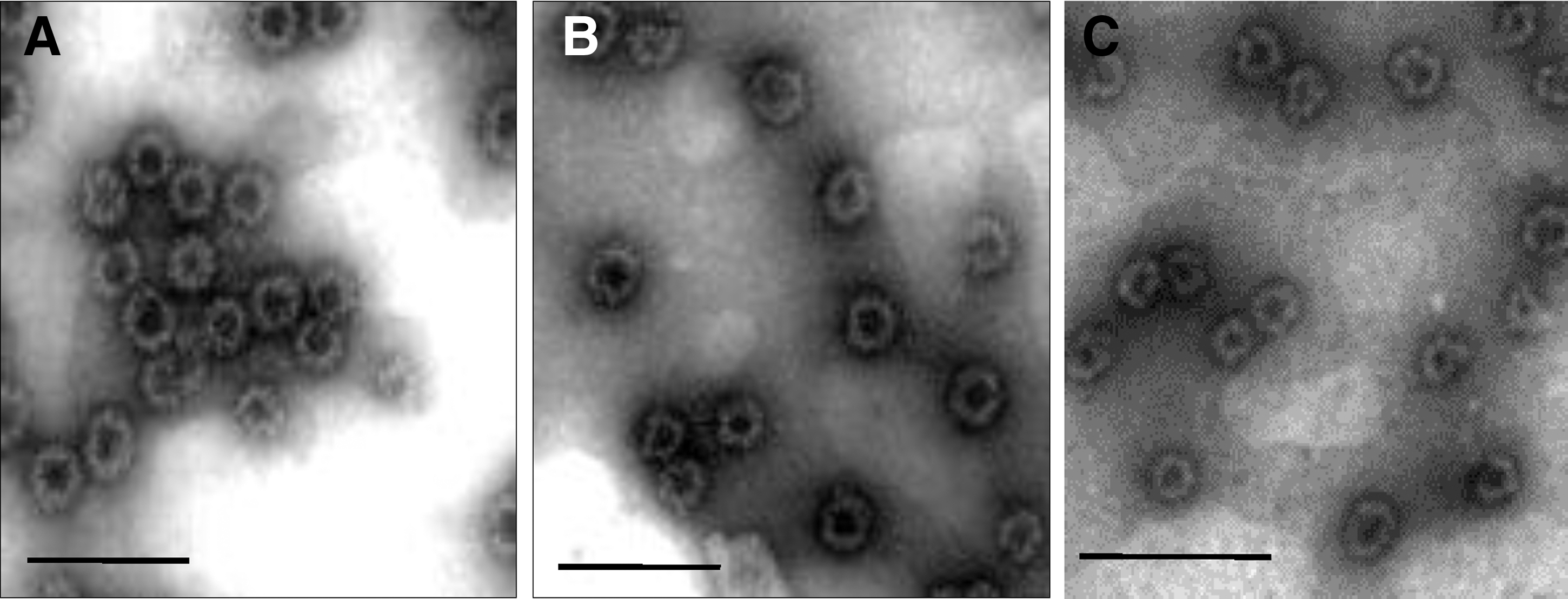
Electron microscopy of sucrose gradient centrifugation-purified noroviruses virus-like particles stained with 1% phosphotungstic acid, pH 7.0.
Farmington-Hill strain is the GII-4-2002 variant.
Seroprevalence of three different NoV strains in a Korean population
The prevalence of serum antibodies to the predominant strains of NoV in Korea as a factor of age is shown in Fig. 3. The overall seroprevalence of immunoglobulin G antibodies against NoV strains GI-4, GII-3, and GII-4 in the 346 serum specimens was 84.1%, 76.3%, and 94.5%, respectively. Seroprevalence was 60.0–85% in children aged <11 months. A rapid decrease in seroprevalence occurred after birth, with the lowest levels (57.1% for GI-4, 47.6% for GII-3, and 66.7% for GII-4) observed in the 12–23-month age group. A sharp increase in seroprevalence occurred in early childhood, reaching 64.6% for GI-4, 68.8% for GII-3, and 91.7% for GII-4 in the 2–5-year age group. The seroprevalence of all three strains/genotypes exceeded 80% in subjects over 20 years. In all age groups, a statistically significant difference was observed in the seroprevalence of different NoV genotypes, with immunity against GII-3 NoV significantly less frequent and immunity against GII-4 NoV significantly more frequent than immunity against the other two genotypes (p<0.01). Among 346 tested sera, 238 sera have detectible antibodies against all kinds of NoV strains used in this study and just 23 sera have detectable antibodies against single stains of NoV (19 for GII-4, two for GII-3, and two for GI-4 respectively). Among the single-strain reactive sera, 11 of 19 in the GII-4 reactive sera and all single reactive sera for GI-4 and GII-3 were collected from enrolled persons under 10 years old.
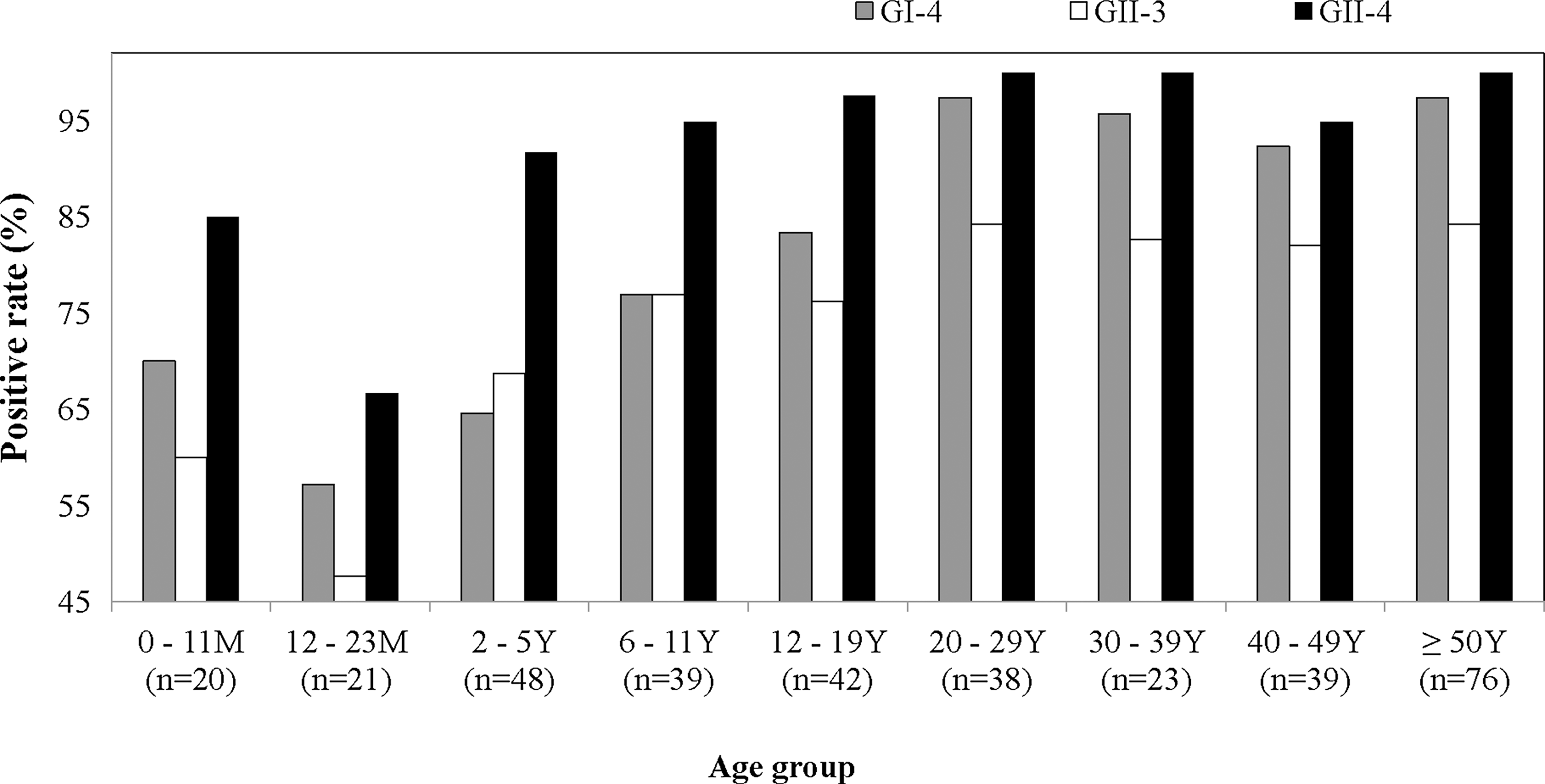
The prevalence of seroresponse to baculovirus-expressed recombinant noroviruses capsid proteins of three different genotypes in Korea. n, number of tested serum samples.
Discussion
Studies of NoV have been severely restricted due to the absence of an established viral cultivation system using cell culture or a small-animal model. VLP expressed in eukaryotic expression systems such as yeast or insect cells are alternative antigens that can be used in lieu of native NoV capsid proteins in studies of immune response against NoV infection (Jiang et al., 1992; Green et al., 1993). The present study describes the expression of recombinant capsid antigens representing three major NoV genotypes with NoV-specific morphology and investigation of the seroprevalence of these NoV strains in Korea by using VLP antigens.
All studies on host–pathogen interactions have come from human challenge and epidemiological investigations (Graham et al., 1994; Rockx et al., 2005; Atmar et al., 2008). According to seroepidemiological studies conducted worldwide, the rate of seropositivity to NoV varies by location, and high rates of seropositivity have been observed for NoV strains associated with recent epidemics. In Hokkaido, Japan, seropositivity for GI-1 and GII-3 has been reported to be 82% and 88%, respectively (Honma et al., 1998). In China, the rate of seropositivity was 89% for GI-1 and 91% for GII-3 (Jing et al., 2000). In Ga-Rankuwa, South Africa, the rate of seropositivity was 96% for GI-1 and 99% for GII-3 (Smit et al., 1997). In Italy, the rate of seropositivity was 51% for GI-2 and 91.2% for GII-4 (Pelosi et al., 1999). Our findings concerning the prevalence of GI-4, GII-3, and GII-4 in the present study are in line with these previous reports.
In this investigation, the changing pattern of seroprevalence according to age was similar in all assessed strains of NoV. Immunity against NoV after birth was higher than in young children aged from 12 to 24 months, and seroprevalence steeply increased from the age of 5 years. The high levels of maternal antibodies observed in newborns indicate a high exposure to these strains in adult women and a high frequency of primary infection in early childhood.
The comparison of the seroprevalence of antibodies to three VLP belonging to different genotypes revealed that infection with GII-4 is more prevalent and occurs earlier than the other two genotypes GI-4 and GII-3, which has been confirmed by several previous investigations. Interestingly, although the nationwide prevalence of GII-3 infection is higher than that of GI-4 in both sporadic cases and epidemics in Korea, GI-4 and GII-4 were more predominant strains observed in the present study, and the incidence of infection by GII-3 was very low in the same region of Korea from 2002 to 2004 (data not shown). Higher exposure to specific strains, including GI-4 and GII-4, may induce a higher prevalence of immunity against these two dominant strains. The seroprevalence to the GI-4 strain remained high even though a very low incidence of infection by GI-4 occurred in the same region. This finding may be the result of cross-reactivity between genotypes due to shared epitopes in the capsid protein. We determined cross-reactivities of antisera raised against three genotype-specific VLPs. These results indicated that GI-4 antisera cross-reacted weakly with other genotypes, but GII-3 and GII-4 antisera showed genotype-specific reactivities (Table S1; Supplementary Data is available online at
Worldwide NoV epidemics have been associated with the predominant genotype GII-4 and have been attributed to the emergence of new GII-4 variants every 1–2 years (Noel et al., 1999; Bull et al., 2006; Siebenga et al., 2007). Multiple recent reports have concluded that the major capsid proteins of GII-4 strains are evolving rapidly, resulting in new epidemic strains with altered antigenicity. The majority of these changes are occurring within the surface exposed P2 subdomian. Surrogate neutralization assays using both sera collected from human GII-4 outbreaks and from NoV-immunized mice suggest that potential neutralizing epitopes are not conserved among GII-4 NoVs (Lindesmith et al., 2011). This antigenic variation and accompanying host immune evasion may contribute to GII-4 persistence in the human population. The high seropositivity to the GII-4 strain may reflect a recent epidemic of GII-4 NoV infection, and emerging GII-4 strains may begin circulation toward the end of an epidemic, as neutralization assays help to identify influenza virus vaccine candidates (Lindesmith et al., 2008; Cannon et al., 2009).
Through this study, recombinant NoV capsid proteins were established as valuable candidates to supply antigens for the development of vaccine and diagnostic antibodies, which, due to the limited number of known NoV self-assembled VLP, may be an important development in the study of NoV. This study is the first report on the prevalence of infection with three dominant NoV genotypes in Korea and has established basic epidemiological information valuable to the understanding of NoV infections in Korean populations. Prospective investigations of dominant genotypes and the serology of new strains or variants will be needed increasingly to predict the spread of NoV infection and aid in vaccine development.
Footnotes
Acknowledgment
We thank the staffs at five heath care centers in Jeollanam-do for collecting serum specimens. This study was supported by a grant from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare & Family Affairs, Republic of Korea (A085119).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.