
Abstract
Broad-spectrum cephalosporin (BSC) resistance has increased in Escherichia coli isolates from broiler chickens in Japan since 2004. The purpose of this study was to understand the epidemiology of BSC-resistant E. coli in livestock animals. Among 3274 E. coli isolates from 1767 feces of apparently healthy animals on 1767 farms between 2004 and 2009, 118 ceftiofur (CTF)-resistant isolates (CTF MIC ≥4 μg/mL) were identified on 74 farms. After elimination of apparently clonal isolates from a single animal, 75 selected CTF-resistant isolates (62 isolates from 61 broiler chickens, 10 isolates from 10 layer chickens, two isolates from two cows, and one isolate from a pig) were characterized. The bla CMY-2 gene was most frequently detected in 50 isolates, followed by bla CTX-M (CTX-M-2: six isolates; CTX-M-14: four isolates; CTX-M-25: two isolates; CTX-M-1: one isolate) and bla SHV (SHV-12: seven isolates; SHV-2, SHV-2a, SHV-5: one isolate each). In particular, 42 of 62 broiler chicken isolates harbored bla CMY-2. Pulsed-field gel electrophoresis analyses using XbaI revealed divergent profiles among the BSC-resistant isolates. The incompatibility groups of bla CMY-2 plasmids from 34 of the 42 broiler chicken isolates belonged to IncIγ (10 isolates), IncA/C (nine isolates), IncB/O (seven isolates) and IncI1 (six isolates), or were nontypeable (two isolates). Co-transmission of resistance to non-β-lactam antibiotics was observed in transconjugants with IncA/C plasmids, but not with IncI1, IncIγ, and IncB/O plasmids except for one isolate with IncB/O. Our findings suggest that the bla CMY-2 gene is a key player in BSC-resistant E. coli isolates and that coselection is unlikely to be associated with the abundance of bla CMY-2 plasmids, except for IncA/C plasmids.
Introduction
T
In Gram-negative bacteria, BSC resistance has been associated with the production of extended-spectrum β-lactamases and AmpC β-lactamases on transferable plasmids (Li et al., 2007). Although the phylogenetic types of E. coli differ between humans and animals (Maynard et al., 2004), common replicon types of plasmids encoding β-lactamase genes were found in E. coli isolates from humans and food-producing animals (Carattoli, 2009). Moreover, transfer of AmpC β-lactamase plasmids between E. coli and Salmonella among humans and animals has been documented (Winokur et al., 2001). Characteristics of the plasmids in E. coli are important for analyzing transmission of resistant bacteria and/or resistance genes from food animals to humans via the food chain. The purpose of this study was to examine the appearance and characteristics of cephalosporin resistance genes and plasmids in BSC-resistant E. coli from livestock animals.
Materials and Methods
Bacterial strains
Data for 3274 E. coli isolates (1095 isolates from 582 cattle farms, 802 isolates from 433 pig farms, 699 isolates from 392 layer chicken farms, and 678 isolates from 360 broiler chicken farms) were collected in the Japanese Veterinary Antimicrobial Resistance Monitoring program from 2004 to 2009. Detailed methods for the sample collection and bacterial isolation were described previously (National Veterinary Assay Laboratory, 2009). The 47 prefectures were divided into four groups between 2004 and 2007 (11 or 12 prefectures per year) and divided into two groups after 2008 (23 or 24 prefectures per year), selected evenly on the basis of the geographical differences from northern to southern areas. Freshly voided fecal samples from healthy cows, pigs, broiler chickens, and layer chickens were collected from six animals of each species at the different farms in each prefecture during 2004–2007. They were collected from about six healthy cows, two pigs, two broiler chickens, and two layer chickens after 2008.
Antimicrobial susceptibility testing
The minimum inhibitory concentrations were determined using a commercially available broth microdilution test (Eiken Co. Ltd., Tokyo, Japan) according to the manufacturer's instruction. To confirm the quality of the products, Staphylococcus aureus ATCC 29213, Enterococcus faecalis ATCC29212, E. coli ATCC 25922, and Pseudomonas aeruginosa ATCC 27853 were used for quality control following the guidelines of the Committee for Clinical and Laboratory Standards Institute (Clinical and Laboratory Standards Institute, 2008). The following antimicrobial agents were tested: ampicillin (AMP, range of antimicrobial dilution: 1–128 μg/mL), piperacillin (PIP, 2–64), cefazolin (CFZ, 1–128), cefpodoxime (CEF, 1–32), cefotaxime (CTX, 0.5–64), ceftazidime (CAZ, 1–64), aztreonam (AZT, 4–16), imipenem (IMI, 1–32), tetracycline (TET, 0.5–64), gentamicin (GEN, 0.5–64), kanamycin (KAN, 1–128), chloramphenicol (CHL, 1–128), nalidixic acid (NAL, 1–128), ciprofloxacin (CIP, 0.03–4), and trimethoprim/sulfamethoxazole (SXT, 2.38/0.12–152/8). Resistance phenotypes were determined following the Clinical and Laboratory Standards Institute standards (Clinical and Laboratory Standards Institute, 2011).
A double-disk synergy test for detection of extended-spectrum β-lactamase was performed using clavulanate/amoxicillin and CTX, CAZ, CEF, or AZT disks (Nissui Pharmaceutical Co. Ltd., Tokyo, Japan) as previously described (Kojima et al., 2005).
Determination of β-lactamase genes
The presence of genes encoding TEM and SHV β-lactamases was determined by polymerase chain reaction (PCR) using specific primers as previously described (Kojima et al., 2005). Multiplex PCR was applied to the detection of the AmpC genes, groups ACC, FOX, MOX, CIT, and EBC (Dallenne et al., 2010), and the CTX-M β-lactamase genes as described elsewhere (Xu et al., 2005). In samples positive for the CIT group, sequence analyses were performed using primer pairs for amplification of the full-length bla CMY-2 gene (Kojima et al., 2005). The subtypes of the CTX-M β-lactamases were determined using primer pairs as follows: CTX-MF (5′-GACTA TTCATGTTGTTGTTATTTC-3′) and CTX-MR (5′-TTACAA ACCGTTGGTGACG-3′) for CTX-M-group1 (Mena et al., 2006); bla CTX-M-2F (5′-ATGATGACTCAGAGCATTCG-3′) and bla CTX-M-2R (5′-TCAGAAACCGTGGGTTACGA-3′) for CTX-M-group2 (Kojima et al., 2005); CTXM825F (5′-CGCTTTGC CATGTGCAGCACC-3′) (Pitout et al., 2004) and CTXM20 (5′-ATAACCGTCGGTGACAATT-3′) for CTX-M-group8 and 25/26 (Xu et al., 2005). Sequencing reactions were performed directly on both strands of the PCR products. The DNA alignments and deduced amino acid sequences were analyzed using the BLAST program (National Center for Biotechnology Information, Bethesda, MD).
Analysis of the chromosomal AmpC promoter region
Sequencing of the chromosomal AmpC promoter region was performed in CTF-resistant isolates, which were confirmed to be negative for the presence of β-lactamase genes by PCR, as previously described (Kojima et al., 2005).
Pulsed-field gel electrophoresis (PFGE) and phylogenetic analysis
CTF-resistant isolates were typed by PFGE using the Xba I restriction enzyme according to the CDC PulseNet protocol (Hunter et al., 2005). Electrophoresis was performed using a CHEF-DR III System (Bio-Rad Laboratories, Hercules, CA) with one phase from 2.2 to 63.8 s at 180 V for 19 h. The PFGE profiles were analyzed using Molecular Analyst Fingerprinting Plus software version 1.6 (Bio-Rad Laboratories). Similarity and diversity were assessed using the Dice coefficient with a 1.0% band position tolerance and 0% optimization settings. The phylogenetic groups of the isolates were determined by multiplex PCR as described by Clermont et al. (Clermont et al., 2000).
Conjugation and incompatibility grouping of plasmids
Plasmids were mated to E. coli DH5α by the broth-mating method (Usui et al., 2012). Briefly, overnight cultures of donor (20 μL) and recipient (20 μL) bacteria were mixed with 160 μL of fresh LB broth and incubated at 35°C overnight. Transconjugants were selected on Mueller-Hinton agar supplemented with 50 μg/mL rifampicin (Sigma-Aldrich, St. Louis, MO) and 32 μg/mL CFZ. Plasmid incompatibility grouping was performed using transconjugants and a PCR-based replicon typing method (Carattoli et al., 2005). PCR-restriction fragment length polymorphism (RFLP) analyses were applied to distinguish between IncI1 and IncIγ. Briefly, PCR amplification was performed using primers for IncI (forward: 5′-GAGCAAAGCGGCGAAAGCCG-3′; reverse: 5′-GCTCTT CGGGTGCTAACTGACTCC-3′), followed by 35 cycles of 94°C for 30 s, 59°C for 30 s, and 72°C for 30 s. The PCR product was 332 bp. When the PCR products of IncI1 were digested at 140–144 (136–145, 5′-ATGG
Plasmid DNA preparation and Southern blotting
Plasmid DNA was purified from the original strains and transconjugants by a modified alkaline lysis method (Kado and Liu, 1981). Chromosomal or plasmid DNA in agarose gels was transferred to N+ Hybond nylon membranes (GE Healthcare, Buckinghamshire, UK). DNA of bla CMY-2, IncI, IncA/C, and IncB/O was amplified with specific primer sets and labeled using a PCR DIG probe synthesis kit (Roche Diagnostics, Mannheim, Germany). Hybridization, washing, and detection were performed using a DIG Easy Hyb, DIG Wash and Block buffer set and DIG enzymatic detection kits (all from Roche Diagnostics), respectively, according to the manufacturer's instructions.
Results
Antimicrobial susceptibility
CTF resistance (CTF minimum inhibitory concentration ≥4 μg/mL) was found in 108 of 3274 (3.3%) isolates from food-producing animals in the Japanese Veterinary Antimicrobial Resistance Monitoring program between 2004 and 2009. The incidence of CTF resistance was 13.0% (89/678) in broiler chicken isolates in 2004–2009. In the other animals, CTF resistance was found in 0.2% (2/1095) of bovine isolates, 0.2% (2/802) of porcine isolates, and 2.1% (15/699) of layer chicken isolates between 2004 and 2009. In the present study, if two CTF-resistant isolates from an animal exhibited the same antimicrobial resistance profile and carried the same β-lactamase genes, they were represented as a single isolate. A total of 75 isolates (62 isolates from 61 broiler chickens, 10 isolates from 10 layer chickens, two isolates from two cows, and one isolate from a pig) were used in further analyses. There were two isolates included from one broiler chicken because each of the isolates contained a different resistant gene (bla CMY-2 and bla CTX-M-2, respectively).
All of the 75 CTF-resistant E. coli isolates exhibited resistance to AMP, CFZ, and CEF, but were susceptible to IMI. AZT resistance was found in 17 CTF-resistant E. coli isolates (22.7%), while resistances to CTX and CAZ were found in 63 (84.0%) and 40 isolates (53.3%), respectively. Except for resistance to β-lactam antibiotics, TET resistance was frequently found in 53 CTF-resistant E. coli isolates (72.0%). The number of isolates resistant to the remaining non-β-lactam antimicrobials tested was less than 50% (Table 1).
Resistance breakpoint (μg/mL−1).
AMP, ampicillin; PIP, piperacillin; CFZ, cefazolin; CPD, cefpodoxime; CTX, cefotaxime; CAZ, ceftazidime; ATM, Aztreonam; IPM, imipenem; TET, tetracycline; GEN, gentamicin; KAN, kanamycin; CHL: chloramphenicol; NAL, nalidixic acid; CIP, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole.
Detection of β-lactamase genes
Of 62 CTF-resistant isolates from broiler chickens, 42 isolates harbored bla CMY-2, 11 harbored bla CTX-M, and eight harbored bla SHV (Table 1). In the broiler chicken isolates, various types of extended-spectrum β-lactamase genes were identified. For the CTX-M enzymes, bla CTX-M-2, bla CTX-M-14, and bla CTX-M-25 were found in the broiler chicken isolates. For the SHV enzymes, bla SHV-2, bla SHV-2a, bla SHV-5, and bla SHV-12 were observed. Regarding the isolates from the other animals, bla CMY-2, bla CTX-M, and bla SHV were found in seven, two, and one layer chicken isolates resistant to CTF, respectively. bla SHV-12 was found in one bovine isolate and bla CMY-2 was found in one porcine isolate.
Detection of mutations in the AmpC promoter region
In two β-lactamase gene-negative CTF-resistant E. coli isolates, one broiler chicken isolate had mutations at bases −88, −82, −18, −1, and +58, and one bovine isolate had mutations at bases −88, −82, −42, −18, −1, and +58 in the AmpC promoter region.
Genetic relatedness of CTF-resistant isolates from food-producing animals
Of the 75 isolates, only 66 isolates were included in the PFGE analyses because nine isolates exhibited smear images. The PFGE analyses with Xba I digestion showed high diversity of the PFGE profiles among the CTF-resistant isolates. Using the unweighted pair group method with arithmetic mean, a low similarity (<80%) was observed in all of the isolates with the exception of two broiler chicken isolates (Fig. 1). There were no detectable relationships between strains according to their animal species, regions of isolation, years of isolation or β-lactamase gene types. Phylogenetic grouping revealed that 31 isolates belonged to group A, 20 belonged to group B1, and 23 belonged to group D (Table 1).
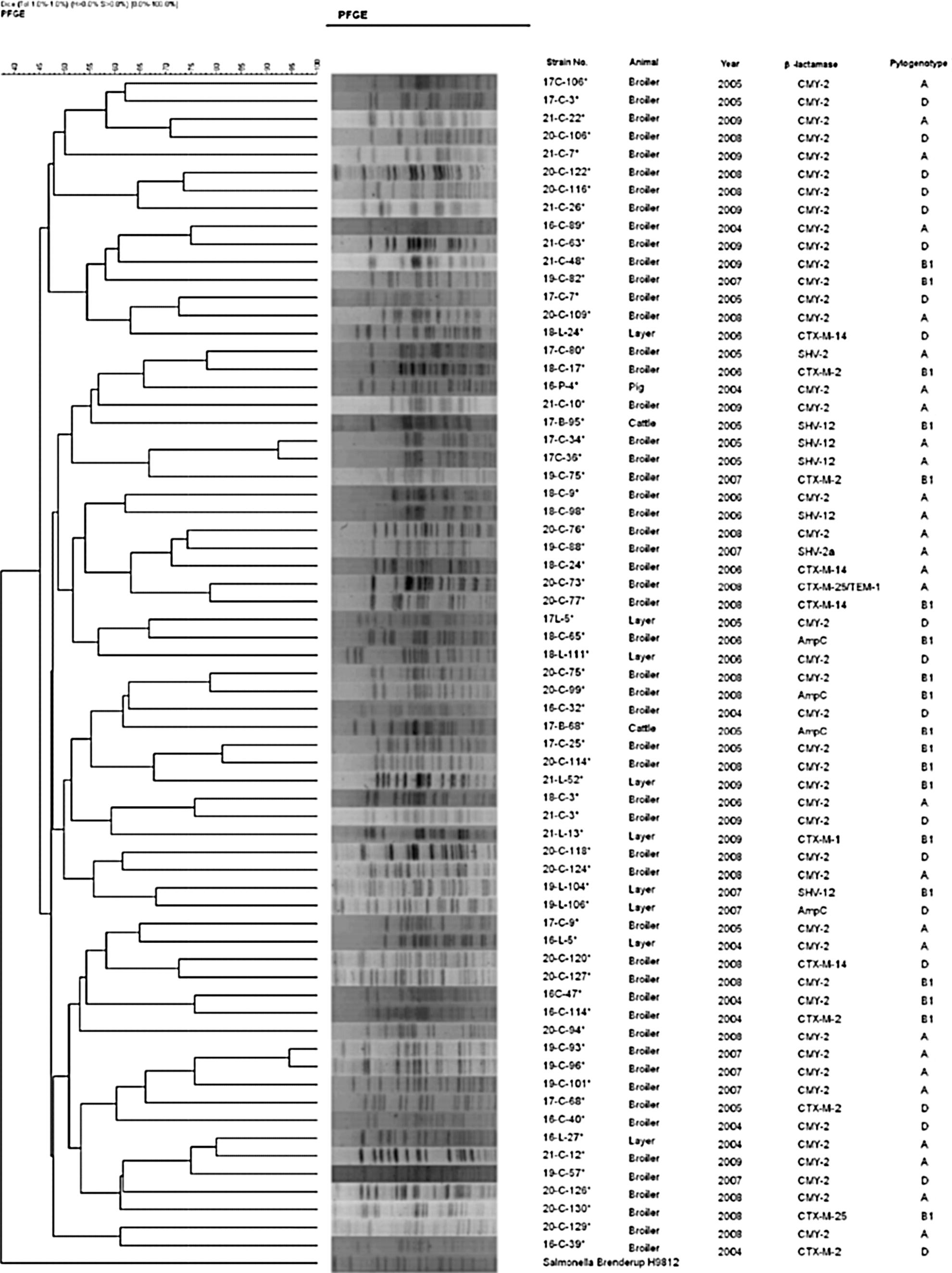
Pulsed-field gel electrophoresis (PFGE) profiles among ceftiofur-resistant Escherichia coli isolates from food-producing animals in Japan. A DNA molecular weight marker derived from Salmonella Braenderup strain H9812 was used as a control.
Characteristics of conjugated plasmids encoding CMY-2 in broiler chicken isolates
Thirty-four transconjugants (82.9%) were obtained from 42 bla CMY-2-positive E. coli. Replicon typing and RFLP analyses using the transconjugants revealed that 32 (94.1%) of the plasmids harboring bla CMY-2 belonged to four distinct incompatibility groups of IncIγ (29.4%), IncA/C (26.4%), IncB/O (20.5%), and IncI1 (17.6%), while the rest were nontypeable (5.9%). Multiple incompatibility types of transferable plasmids were observed for every year in the periods studied (Table 2).
TET, tetracycline; GEN, gentamicin; KAN, kanamycin; CHL, chloramphenicol; NAL, nalidixic acid; CIP, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole.
Transfer of resistance to antimicrobials other than β-lactam antibiotics was observed in the transconjugants with IncA/C (Table 2), of which four showed resistance to TET-CHL, two showed resistance to TET-SXT, two showed resistance to TET-CHL-SXT, and one showed resistance to GEN-KAN-TET-CHL. Of seven transconjugants with IncB/O plasmids, one exhibited resistance to TET-ST and the remaining six were susceptible. All of the transconjugants with IncI1-Iγ plasmids were susceptible to all the non-β-lactam antimicrobials tested.
Discussion
The present study has shown that the bla CMY-2 gene is a key player in BSC-resistant E. coli isolates from food-producing animals, especially broiler chickens. In Japan, the presence of bla CMY-2 has been consistently observed in E. coli from apparently healthy broiler chickens (Kojima et al., 2009) since its first isolation in 1999 (Kojima et al., 2005). The presence of the gene was also detected in layer chicken isolates after 2003 (Kojima et al., 2009) and in porcine isolates after 2004 in this study. The percentages of BSC resistance remain low in E. coli from cattle, pigs, and layer chickens (National Veterinary Assay Laboratory, 2009). In the present study, the BSC-resistant isolates from broiler chickens were classified almost equally into three groups, A, B1, and D. Group A was frequently found in most of CFZ-resistant E. coli from poultry affected with colibacillosis in Japan (72.4%, 21/29) (Asai et al., 2011). In addition, group B2, which is frequently found in human pathogenic E. coli, was not prevalent in the isolates tested (Maynard et al., 2004). PFGE analyses revealed great diversity among the E. coli isolates from livestock in this study. Other study showed different PFGE profiles of BSC-resistant E. coli isolates from broiler chickens on several farms (Hiroi et al., 2012). In Japan, although clonal spread of Salmonella was observed in broiler chickens (Asai et al., 2009), the genetic relatedness of CMY-2-producing E. coli in food-producing animals was low.
Various replicon types of plasmids mediating BSC resistance are globally distributed in Enterobacteriaceae (Carattoli, 2009). The present study showed that four replicon types—IncI1, IncIγ, IncA/C, and IncB/O—of transferable plasmids encoding bla CMY-2 were responsible for the appearance of BSC-resistant E. coli among broiler chickens. The IncA/C, IncI1, and IncIγ plasmids encoding bla CMY-2 have been prevalent among a wide range of Enterobacteriaceae isolated from humans and animals worldwide (Antunes et al., 2010; Carattoli, 2009; Folster et al., 2011; Mataseje et al., 2009; Poole et al., 2009). Inc B/O plasmids were previously reported in Salmonella isolated from animals in the United States (Lindsey et al., 2009; Poole et al., 2009) and among sulphonamide-resistant E. coli isolated from humans and pigs in Denmark (Wu et al., 2010), but have never been associated with CMY-2 prior to this study.
The emergence and prevalence of antimicrobial resistance in bacteria are likely to be linked to therapeutic antimicrobial use in food-producing animals through cross-resistance and co-resistance (Harada and Asai, 2010). In Japan, penicillin antibiotics, such as AMP, amoxicillin, and penicillin G, are approved and used for the treatment of poultry diseases such as colibacillosis, staphylococcosis, and necrotic enteritis. Between 2001 and 2007, the incidence of AMP resistance remained at stable levels between 40.3% and 53.3% in E. coli isolates from broiler chickens in Japan (National Veterinary Assay Laboratory, 2009). Thus, use of penicillin antibiotics does not act as a selective force. Dutil et al. (2010) indicated that the observed increases in cephalosporin resistance in Salmonella Heidelberg and E. coli isolates from broiler chickens and retail meats in Canada were related to off-label CTF use in hatcheries. In Japan, although off-label use of CTF had been performed for in ovo vaccination or vaccination of 1-day-old chicks in some hatcheries, the off-label use of CTF for poultry was voluntarily banned around March 2012. Coselection using non-β-lactam antibiotics may contribute to the occurrence of specific plasmids in bacteria (Harada and Asai, 2010). IncA/C plasmids encoding AmpC β-lactamases confer resistance to non-β-lactam antibiotics, while IncI1-Iγ plasmids confer resistance to β-lactam antibiotics (Folster et al., 2011; Mataseje et al., 2009; Poole et al., 2009). In the present study, resistance to TET, CHL, and SXT was transferred with β-lactam resistance in BSC-resistant E. coli harboring bla CMY-2-encoding IncA/C plasmids with divergent RFLP profiles. TET antibiotics are most frequently used for treatment of diseases in poultry, and phenicols (florfenicol and thiamphenicol) and trimethoprim-sulfonamide combination drugs are also used in poultry (Asai et al., 2005; Harada and Asai, 2010). Continuous monitoring may help to evaluate the effects of CTF use and the contribution of coselection to the appearance of bla CMY-2 plasmids in E. coli from broiler chickens.
Conclusions
The present study has shown that dissemination of multiple types of plasmids encoding bla CMY-2 is involved in the increase in BSC resistance in E. coli in broiler chickens in Japan. Of the four types of plasmids detected, IncI1 and IncA/C plasmids are common in several countries, while IncIγ and IncB/O plasmids are rare. Continuous investigations at the national level for BSC-resistant E. coli in food-producing animals remain a high priority to clarify the contribution of antimicrobial use to the dissemination and increase in plasmids encoding bla CMY-2.
Footnotes
Acknowledgments
We thank the staff of the Livestock Hygiene Service Centers across Japan for providing the E. coli isolates. This work was supported in part by a grant-in-aid from the Japanese Ministry of Health, Labour and Welfare (H21-Shokuhin-Ippan-013).
Disclosure Statement
No competing financial interests exist.