
Abstract
Campylobacter jejuni is one of the most important causes of foodborne gastroenteritis. Chickens are considered a reservoir host of C. jejuni, and epidemiological studies have shown that contaminated chicken meat is a primary source of human infection. The objective of this study was to produce chicken egg-yolk-derived antibody (IgY) against the five C. jejuni colonization-associated proteins or CAPs (CadF, FlaA, MOMP, FlpA, and CmeC). Recombinant C. jejuni CAPs were expressed in Escherichia coli and were purified by affinity chromatography. Specific-pathogen-free laying hens were hyperimmunized with each recombinant CAP to induce production of α-CAP-specific IgY. Egg yolks were collected from immunized and nonimmunized hens and were lyophilized to obtain egg-yolk powder (EYP) with or without α-C. jejuni CAP-specific IgY. IgY was purified from EYP, and the antibody response in serum and egg yolk was tested by indirect enzyme-linked immunosorbent assay. The α-C. jejuni CAP-specific IgY levels were significantly (p<0.05) higher in both serum and EYP obtained from immunized hens as compared with the nonimmunized hens. Each α-C. jejuni CAP-specific IgY reacted with the C. jejuni cells and recombinant CAPs as detected by immunofluorescence microscopy and Western blot assays, respectively. We also show that α-CadF, α-MOMP, and α-CmeC IgY significantly reduced adherence of C. jejuni to the chicken hepatocellular carcinoma (LMH) cells, suggesting that these α-C. jejuni CAP-specific IgY may be useful as a passive immunotherapeutic to reduce C. jejuni colonization in chickens.
Introduction
C. jejuni-infected chickens transfer anti-C. jejuni maternal antibodies (IgY) to the new hatchlings that protect against C. jejuni colonization until 2 weeks of age (Sahin et al., 2003). More importantly, previous studies have shown that these maternal IgY are specifically reactive against C. jejuni colonization-associated proteins (CAPs) including CadF, FlaA, FlaB, PorA (MOMP), and CmeC (Sahin et al., 2003; Shoaf-Sweeney et al., 2008; Cawthraw and Newell, 2010). The C. jejuni CAPs are known to contribute to host-cell adherence (adhesins), motility, and survival within the host. The adhesins such as CadF (Campylobacter adhesion to fibronectin) (Ziprin et al., 1999) and FlpA (fibronectin-like protein A), (Flanagan et al., 2009) mediate the binding of C. jejuni to fibronectin on epithelial cells in vitro (Monteville et al., 2003; Konkel et al., 2010). Mutation of the cadF and flpA genes results in impaired colonization of C. jejuni in chickens (Flanagan et al., 2009). In C. jejuni, FlaA (major flagellin) and FlaB (minor flagellin) filament proteins are required for maximal motility (Newell et al., 1985; Nuijten et al., 1990; Nachamkin et al., 1993; Guerry, 2007). Mutation of flaA results in significantly reduced intestinal colonization by C. jejuni in chickens (Nachamkin et al., 1993). Finally, membrane-associated proteins such as CmeC and PorA (the major outer membrane protein or MOMP) are also required for colonization of C. jejuni in chickens and mice (Lin et al., 2002; Islam et al., 2010). MOMP is required for binding to cell membrane in cultured human intestinal epithelial cells (Moser et al., 1997). CmeC is part of a multidrug efflux pump (CmeABC) that contributes to bile resistance in C. jejuni colonization in chickens. It is notable that in addition to being highly conserved among different C. jejuni strains, all of the above proteins are exposed on the bacterial surface and therefore potentially available for binding and neutralizing effects of antibodies.
Chicken egg-yolk-derived IgY has been experimentally used in vitro and in vivo to reduce colonization of poultry by pathogens such as Salmonella Enteritidis, Salmonella Typhimurium, Escherichia coli O157:H7, infectious bursal disease virus (Eterradossi et al., 1997; Tsubokura et al., 1997; Lee et al., 2002; Kassaify and Mine, 2004; Chalghoumi et al., 2009a; Chalghoumi et al., 2009b). However, the use of IgY to reduce C. jejuni colonization in chickens remains an underdeveloped strategy. Treatment with antigen-specific IgY is known to significantly reduce host-cell colonization by other foodborne pathogens such as E. coli O157:H7 and Salmonella (Chalghoumi et al., 2009b; Cook et al., 2007; Neri et al., 2011). Therefore, the objective of this study was to produce antigen-specific IgY antibodies against five recombinant-C. jejuni CAPs (CadF, FlaA, CmeC, FlpA, and MOMP) and to test their potential to reduce C. jejuni colonization of host cells.
Materials and Methods
Cloning of C. jejuni CAPs
The genes encoding C. jejuni CAPs were polymerase chain reaction amplified using 50 ng of genomic DNA extracted from C. jejuni NCTC11168 (Parkhill et al., 2000), 250 μM of dNTPs, 10 μM of each primer (Table 1), 2 mM MgSO4, 5 U of PƒuUltra II fusion HS DNA polymerase (Stratagene, USA), and 1X PƒuUltra II reaction buffer. PCR conditions included denaturation at 95°C for 2 min followed by 30 one-step cycles of 95°C for 20 sec, 48°C (porA and flpA), 45°C (cadF and flaA), and 40°C (cmeC) for 20 sec and 72°C for 15 sec with a final extension step for 72°C for 3 min. The PCR products were purified using QIAquick PCR purification kit (Qiagen, USA) and cloned into pET100/D-TOPO vector (Invitrogen).
To obtain directional cloning, each forward primer included a four-base-pair sequence (
Sp, signal peptide.
Expression and purification of recombinant CAPs
The recombinant CAPs (rCadF, rCmeC, rFlaA, rFlpA, and rMOMP) were expressed in E. coli BL21 as N-terminal hexahistidine–glycine-tagged proteins, purified using denaturing or native conditions as described previously (Shah et al., 2008). Proteins were dialyzed using 10 MWCO SnakeSkin dialysis tubing (Thermo Scientific, USA) overnight at 4°C with gentle stirring in phosphate-buffered saline (PBS) (4.3 mM Na2HPO4, 1.4 mM KH2PO4, 2.7 mM KCl, 137 mM NaCl) and the concentration was determined by Micro BCA Protein Assay Kit (Pierce). The purified proteins were visualized by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot as described previously (Shah et al., 2008).
Immunization of chickens
Specific-pathogen-free (SPF) fertile eggs were purchased from Sunrise Farms Inc. (Castskill, NY) and hatched at the university facility followed by housing in C. jejuni–free isolation cages until 35 weeks of age in accordance with the protocols approved by the Washington State University—Institute for Animal Care and Usage Committee (IACUC). C. jejuni–free status of hens was maintained during entire experimental period by culturing cloacal swabs from all hens during the growing period as well as before and after immunization. At 35 weeks, C. jejuni–free hens were distributed into six groups (n=3/group) and were provided ad libitum access to feed and water. A water-in-oil emulsion was prepared by mixing 0.5 mL of PBS containing ∼100 μg of each rCAP with 0.5 mL of Freund's complete adjuvant (Sigma, USA). Each hen was intramuscularly injected at four sites in breast muscles with a total of 1 mL of the emulsion (0.25 mL per site). Booster immunizations were given at 2, 4, and 6 weeks after the first immunization with Freund's incomplete adjuvant (Sigma) emulsified with ∼100 μg of respective rCAP. One group served as nonimmunized control. Eggs were collected daily for 1 week pre-immunization and up to 10 weeks postimmunization, wiped with 70% ethanol, and stored at 4°C until further processing. Blood (2 mL) was collected from the brachial vein of all hens prior to each immunization and at 2-week intervals until the end of the experiment. Serum samples were harvested from the blood and stored at −80°C.
Measurement of serum antibody response
Antibody response was determined by indirect enzyme-linked immunosorbent assay (ELISA) (Sunwoo et al., 1996; Cook et al., 2007). Briefly, 96-well flat-bottom polystyrene plates (Falcon, USA) were coated overnight at 4°C with 1 μg of purified rCAP suspended in 50 mM coating buffer (1.59 g Na2CO3, 2.93 g NaHCO3 in 1 L distilled water, pH 9.6). The plates were blocked for 1 h with 5% skim milk PBS-T followed by addition of 100 μL serum (diluted 1:1000 in 0.1% skim milk PBS-T) for 16 h. After washing, plates were incubated with 100 μL of horseradish peroxidase (HRP)-conjugated donkey-anti-chicken IgY (1:15,000; Gallus Immunotech, Canada) and developed using 100 μL HRP substrate (Bio-Rad) for 15 min. The absorbance was read at 415 nm (BioTek, USA). Six replicates of each serum sample were tested twice. Mean absorbance OD415 values (mean±standard error of the mean [SEM]) for each serum samples were calculated.
Preparation of lyophilized egg-yolk powder (EYP)
Eggs were wiped with 70% ethanol, broken to aseptically separate egg yolk from egg white. Egg yolks were collected in sterile tubes and mixed with equal volume of sterile distilled water followed by freezing at −80°C overnight. Frozen egg yolks were lyophilized for 48 h with vacuum level reaching 133×10−3 mBar and condenser temperature at least −40°C in a Freeze Dry System (Labconco, USA). The EYP was stored at 4°C until further processing.
Purification and quantification of anti-C. jejuni CAP-specific IgY in EYP
The IgY was extracted from each EYP sample by a chloroform extraction method (Hamal et al., 2006; Silim and Venne, 1989). Briefly, 3 mL of sterile PBS were added to 1 g of EYP, vortex mixed, followed by addition of an equal volume of chloroform. The mixture was centrifuged at 4000×g for 30 min and the water-soluble fraction (WSF) was collected. The total protein concentration of the WSF was measured using BradFord assay (Bio-Rad). The purity of IgY preparations was tested by SDS-PAGE (data not shown). The level of anti-C. jejuni CAP-specific IgY against each recombinant CAP and cross-reactivity against C. jejuni whole-cell lysates was determined by Western blot as described below, with the exception that WSF containing IgY was diluted 1:500 (α-MOMP, α-FlaA, and α-FlpA) or 1:50 (α-CadF and α-CmeC) before being used as primary antibody. For the preparation of the whole-cell lysates, C. jejuni pellets were resuspended in PBS and sonicated for 1 min×3 cycles at level 10 with a Misonix cup-horn sonicator (Misonix Inc., USA).
Determination of reactivity of anti-CAP-specific IgY from EYP
The reactivity of the anti-CAP IgY of each hyperimmunized EYP (HEYP) preparations with the respective rCAP was determined by Western blot and with the C. jejuni cells by immunofluorescence microscopy. For Western blot, 0.5 μg of each purified rCAP was transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, USA). The WSF prepared from HEYP or nonimmunized EYP (NEYP) were used as primary antibodies (1:10,000 dilution) and the HRP-conjugated donkey-anti-chicken IgY was used as secondary antibody (1:5000 dilution). For immunofluorescence microscopy, 20 μL of C. jejuni cell suspensions made in PBS were spread onto a clean immunohistochemistry slide and air dried. The smear was heat fixed and the slides were rinsed for 1 min with PBS. The smears were probed with anti-CAP-specific IgY (1:50) and incubated at 37°C for 45 min. Smears were washed twice with PBS followed by probing with 1:200 dilution of fluorescein isothiocyanate–conjugated rabbit anti-chicken IgY (Sigma, USA) at 37°C for 45 min. The smears were washed 10 times with PBS, air dried, and images were captured using a compound fluorescence microscope (Axioscope 2 plus, Zeiss).
Cell adherence assay
Chicken hepatocellular carcinoma cells (LMH) (Kawaguchi et al., 1987) were cultured in Weymouth's media (Sigma) with 10% fetal bovine serum (FBS) (Sigma) in 12-well plates pretreated with 0.1% gelatin for 15 min. Cells were cultured to a density of 2×106 cells per well and incubated for 24 h at 37°C in a humidified, 5% CO2 incubator. C. jejuni cell suspension in Weymouth's media (2×109 CFU/mL) was mixed with either IgY prepared from HEYP or NEYP at a final concentration of 1:10 (antibodies:media) and incubated at 37°C for 30 min. Approximately 1 mL of this suspension was added to each well followed by centrifugation at 500×g for 5 min to promote bacterium–host cell contact. The plates were incubated at 42°C for 2 h. Cells were rinsed three times with PBS to remove non-cell-associated bacteria and lysed with 1 mL 0.5% (v/v) Triton X-100 (Sigma) for 10 min at 42°C. Viable counts of the cell-associated bacteria in the lysate were made on Campy Cefex agar. Data from triplicate samples were averaged and the percent binding for each antibody treatment was calculated by the following formula: cell-bound CFU/inoculum CFU×100.
Data analysis
Data were expressed as means±standard errors. The statistical significance was evaluated by one-factor analysis of variance followed by Tukey's test using Prism5 (GraphPad Inc., USA). P values of<0.05 were considered statistically significant.
Results and Discussion
In this study, we successfully expressed and purified five well-characterized, surface-exposed rC. jejuni CAPs that are not only conserved among C. jejuni strains, but are also required for colonization in chickens (Nachamkin et al., 1993; Moser et al., 1997; Lin et al., 2002; Flanagan et al., 2009). SDS-PAGE analysis revealed an intense single band of expected size for each rCAP (Fig. 1). The identity of each rCAP was also confirmed by immunoblotting with anti-HisG antibody (data not shown). Hyperimmunization of SPF hens with rCAP resulted in the strong serum antibody response as detected by indirect ELISA (Fig. 2). The serum antibody levels at 2, 4, 6, 8, and 10 weeks postimmunization were significantly higher (p<0.001) when compared with the levels prior to immunization. The increase in the activity of serum α-CAP-specific IgY was apparent at 2 weeks after initial immunization, peaked at 3rd or 4th immunization, and then plateaued until the end of the experiment, indicating that all the rCAPs induced a measurable antibody response (Fig. 2). These results corroborate with the previously published reports showing similar trends in the serum antibody response in hens hyper-immunized with whole-cell lysates or purified recombinant proteins (Imberechts et al., 1997; Lee et al., 2000; Shin et al., 2002; Gurtler et al., 2004; Gurtler and Fehlhaber, 2004; Cook et al., 2007; Buckley et al., 2010).
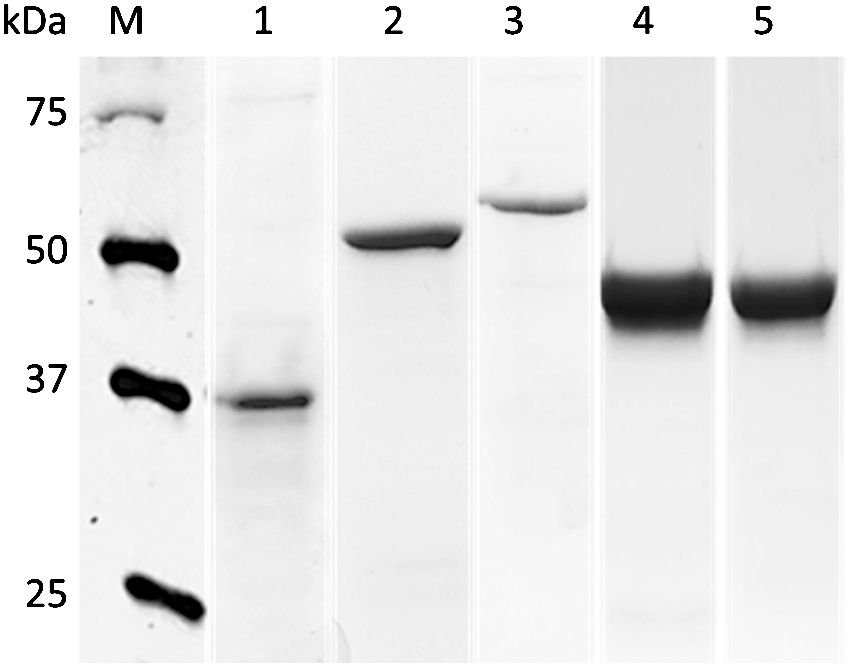
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis stained with Coomassie brilliant blue R-250 showing purified recombinant Campylobacter jejuni colonization-associated proteins. Lanes: M, protein marker (Bio-Rad); 1, CadF (37 kDa); 2, CmeC (55 kDa); 3, FlaA (59 kDa); 4, FlpA (46 kDa) and 5, MOMP (45 kDa).
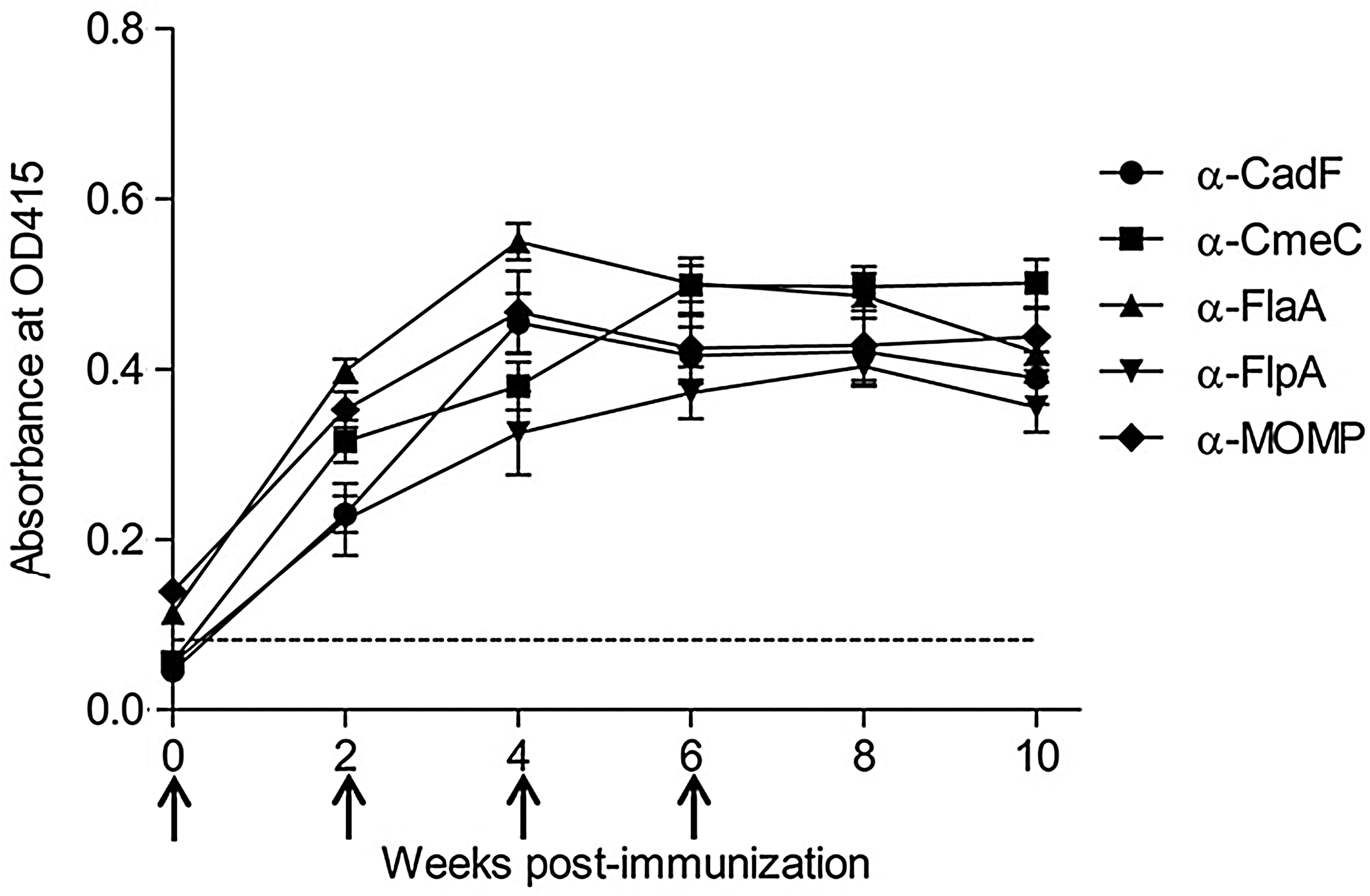
The kinetics of egg-yolk-derived antibody (IgY) response in 1000-fold diluted serum against recombinant Campylobacter jejuni colonization-associated proteins expressed as the enzyme-linked immunosorbent assay value (OD at 415 nm). Each time point represents the mean±standard error of the mean (n=4). The arrows indicate the week of immunization. Dashed line indicates the mean OD415 of nonimmunized sera (negative control). For each time point, IgY titer means differ significantly (p<0.05) from day 0.
It has been reported that the IgY antibody levels in the egg yolk are directly proportional to the IgY levels in the hen's serum (Loeken and Roth, 1983; Hamal et al., 2006). Because all hens immunized with rCAPs showed a peak serum antibody response from the third immunization onwards (Fig. 2), the HEYP collected from the third immunization until the end of the immunization period (i.e., 10 weeks) were pooled for each hen for the determination of reactivity of αCAP-IgY. The level of reactivity of IgY in each of the pooled HEYP against the respective rCAP was significantly higher (p<0.05) as compared with the pre-immunized EYP collected from each group (Fig. 3). We also tested the reactivity of the HEYP with rCAPs by immunoblot assay. For this assay, IgY extracted from the cocktail of all five HEYPs was probed against each of the purified rCAP on the membrane. As expected, each rCAP was identified as a single band (Fig. 4), confirming our previous observation that the HEYP cocktail is enriched with IgY reactive against each of the rCAP. In contrast, IgY extracted from the NEYP did not react with any of the rCAPs. The above results, however, suggest that hens immunized with rCAPs induced a detectable anti-CAP-specific antibody response in both serum and egg yolks and that the hyper-immunized EYP prepared in this study showed strong reactivity with C. jejuni CAPs.
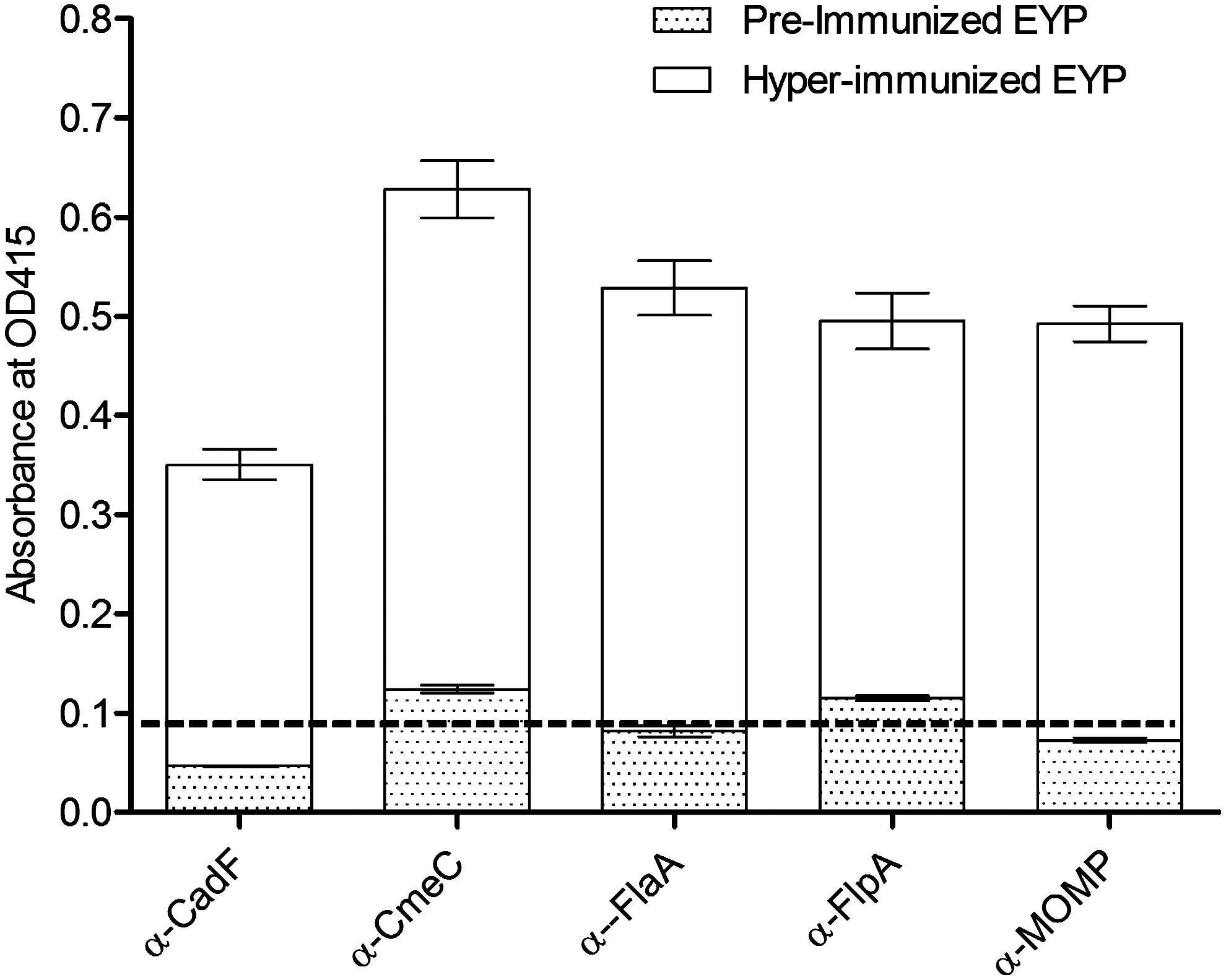
Antibody response in 1024-fold dilution of water-soluble fraction of egg-yolk-derived antibody (IgY) extracted from hyperimmunized egg-yolk powder (EYP, blank bars) against recombinant Campylobacter jejuni colonization-associated proteins (CAPs) and from pre-immunized EYP (dotted bars) using indirect enzyme-linked immunosorbent assay (ELISA). Each bar represents the mean±standard error of the mean (n=4) of the ELISA OD415. Dashed line indicates the mean OD415 of nonimmunized sera (negative control). ELISA showed significantly higher levels of anti-CAP-specific IgY in hyperimmunized EYP compared with the IgY extracted from pre-immunized or nonimmunized EYP (p<0.001).
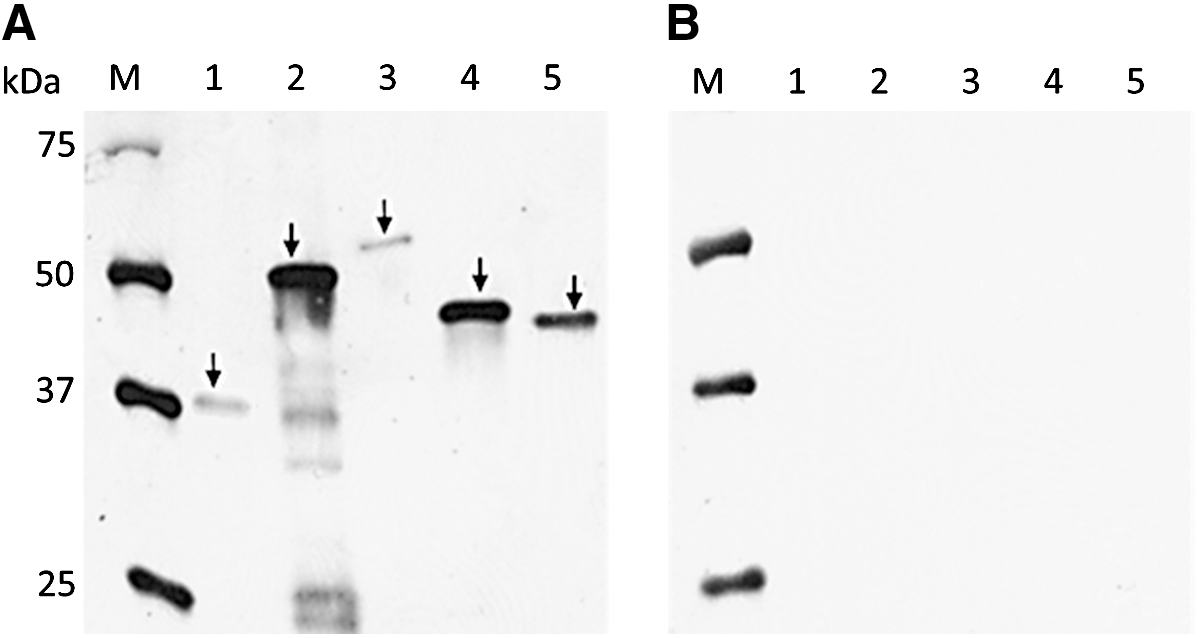
Reactivity of water-soluble fraction (WSF) of egg-yolk-derived antibody (IgY) extracted from hyperimmunized egg-yolk powder (EYP)
It is important to note that none of the SPF hens were exposed to C. jejuni throughout the experimental period. This was also demonstrated by the fact each antibody preparation reacted with specific CAPs with no or negligible reactivity against other proteins from C. jejuni whole cell lysates (Fig. 5). Therefore, we do not expect significant cross-reactivity due to the antibodies produced against natural infection with C. jejuni; however, the possibility of cross-reactivity with nonspecific antibodies directed against other normal-flora bacteria or an adjuvant could not be excluded entirely. We also confirmed the ability of the α-CAP-specific IgY preparation to bind C. jejuni cells by immunofluorescence microscopy. Each α-CAP-specific IgY preparation was able to bind C. jejuni cells as the fluorescence signal was evident in each case (Fig. 6). In contrast, IgY prepared from NEYP did not bind with C. jejuni cells as evidenced from lack of fluorescence (Fig. 6). These results suggest that the α-CAP IgY from each HEYP was bound to C. jejuni cells. Additionally, the α-CAP IgY from each HEYP also showed binding with heterologous strains of C. jejuni, including C. jejuni 81-176 (Hofreuter et al., 2006), RM1221 (Fouts et al., 2005), and F38011 (Larson et al., 2008) (data not shown). The broad-spectrum binding ability of α-CAP IgY to heterologous strains of C. jejuni raises the possibility of usefulness of these IgY preparations as passive immunotherapeutic to reduce C. jejuni colonization in naturally infected chickens.

Reactivity of water-soluble fraction of egg-yolk-derived antibody (IgY) extracted from nonimmunized egg-yolk powder (EYP) (lane 1) and hyperimmunized EYP against whole-cell lysates of Campylobacter jejuni. Lanes 2 (FlaA, 59 kDa), 3 (MOMP, 45 kDa), and 4 (FlpA, 46 kDa) were probed with 1:500 dilution of hyperimmunized EYP against respective recombinant colonization-associated proteins (rCAPs), whereas lanes 5 (CmeC, 55 kDa) and 6 (CadF, 37 kDa faint band) were probed with 1:50 dilution of hyperimmunized EYP against respective rCAPs. Lane M, prestained marker (Bio-Rad).
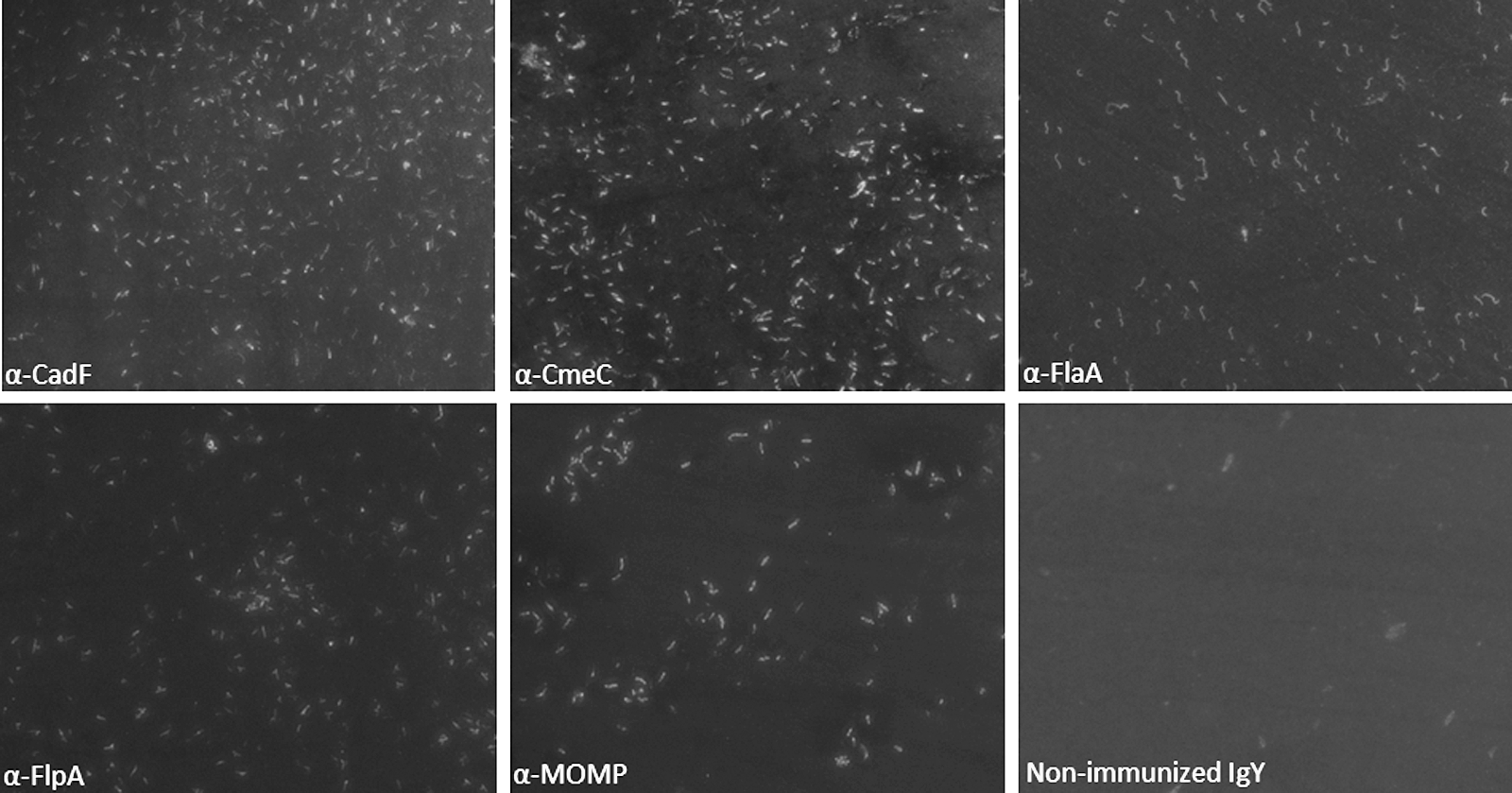
Immunofluorescence micrographs of Campylobacter jejuni–incubated probed with egg-yolk-derived anti-C. jejuni colonization-associated proteins–specific egg-yolk-derived antibody (IgY) and nonimmunized IgY (magnification 63×).
Previous studies using C. jejuni adherence to chicken hepatocellular carcinoma (LMH) or human intestinal epithelial cells as a surrogate model have shown that adherence to the host epithelial cells is requisite for intestinal colonization of this foodborne pathogen (Larson et al., 2008; Flanagan et al., 2009). We hypothesized that egg-yolk-derived antibody (IgY) directed against rCAPs will reduce the host-cell binding of C. jejuni. Consequently, we tested the ability of α-CAP-specific antibodies in reducing C. jejuni binding with in vitro cultured LMH cells. LMH cells were chosen because the majority of the CAPs are known to contribute to binding to LMH cells (Flanagan et al., 2009). The results of LMH cell-binding assay (Fig. 7) revealed that treatment with α-FlaA (19±2% binding) and α-FlpA (19±5% binding) antibodies had little effect on the ability of C. jejuni to bind to the LMH cells when compared with the non-IgY treated infection control (25±2% binding). There was, however, a significant reduction in the binding of C. jejuni when pretreated with α-CadF (11±2% binding), α-MOMP (11±1% binding), and α-CmeC (11±1% binding)-specific IgY (Fig. 7). In contrast, there were no differences between the total number of cell-bound C. jejuni between nontreated infection control (25±2% binding) and C. jejuni pretreated with nonimmunized IgY (25±2% binding). These results indicate that the α-CAP-specific IgY prepared from egg yolk of immunized hens is effective in reducing the binding of C. jejuni to chicken epithelial cells.
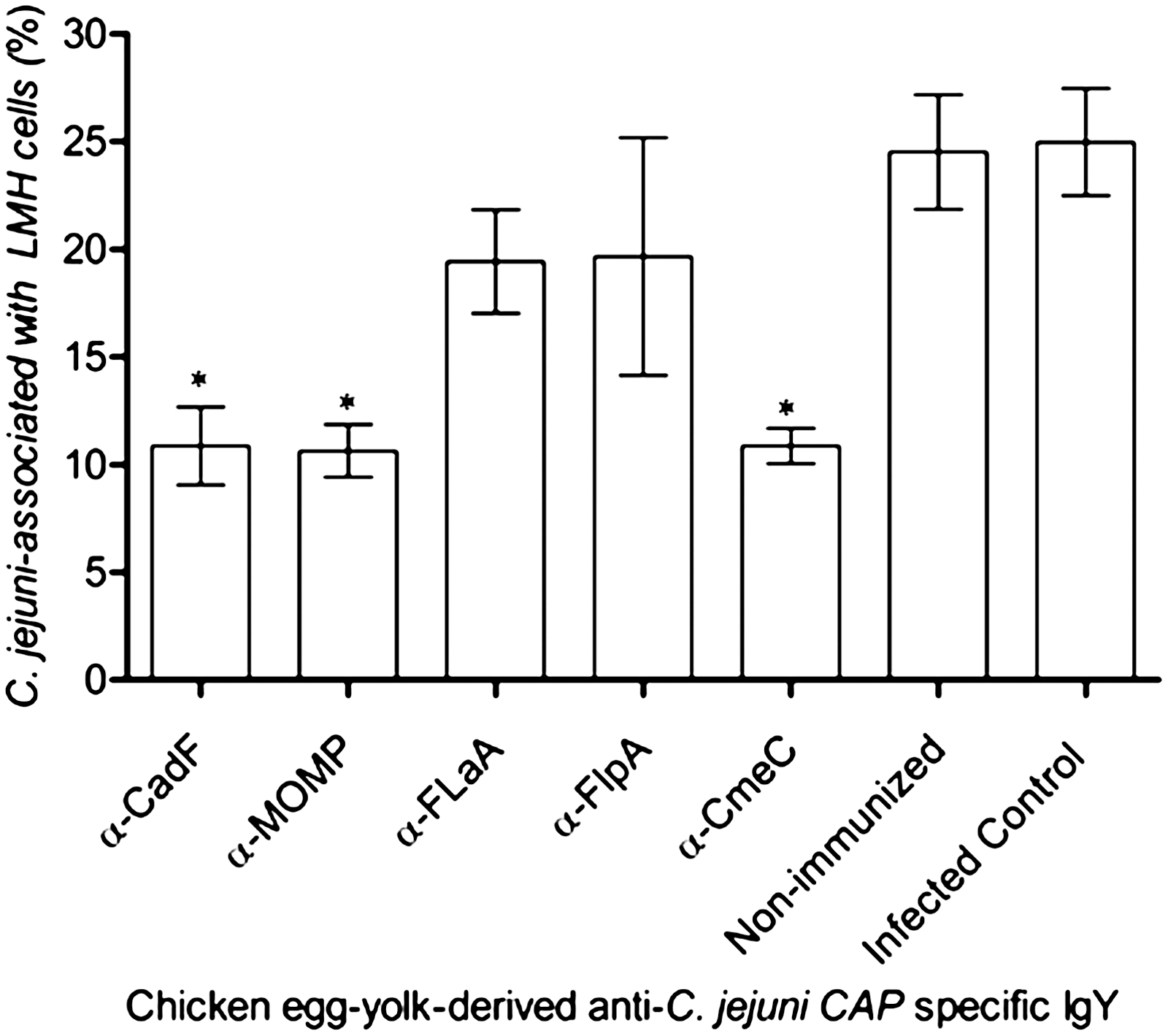
Percentages of Campylobacter jejuni cells associated with chicken hepatocellular carcinoma epithelial cells (LMH) when treated with 10-fold dilution of each α-C. jejuni colonization-associated proteins–specific egg-yolk-derived antibody. Cell adherence assays were performed as described in Materials and Methods. Each bar represents the mean±standard error of the mean (n=3) of C. jejuni cells bound to LMH cells. An asterisk indicates a statistically significant difference (p<0.05) between the C. jejuni–infected control and antibody-treated cells.
Based on the experimental data presented here, it appears that the reduction in C. jejuni adherence to host cells is likely due to the binding of α-CAP-specific IgY to the surface exposed CAPs. Other mechanisms such as inhibition of host-cell colonization by inhibiting motility and/or efficient uptake of nutrients cannot be ruled out. Limited studies have been conducted to test the efficacy of IgY to reduce intestinal colonization of C. jejuni in chickens. Tsubokura et al. (1997) showed that oral administration of a single dose of egg-yolk-derived IgY against C. jejuni whole-cell lysates led to a short-term (12 h post-treatment), 2 log reduction of C. jejuni colonization in chickens. It is possible that the IgY raised against whole-cell lysates contain limiting concentrations of CAP-specific IgY, leading to short-lived protection in the above study (Tsubokura et al., 1997). Our proposed hypothesis is that the short-term treatment using feed supplemented with HEYP enriched with α-CAP-specific IgY will reduce colonization of C. jejuni in chickens. Similar to the principle involved in passive immunity, anti-CAP IgY to the surface-exposed CAPs can potentially inhibit C. jejuni motility, attachment to intestinal cells, and/or efficient uptake of nutrients, thus decreasing the ability of C. jejuni to effectively compete with native microbiota. The antigenic targets described here have a potential for prevention of colonization of C. jejuni at multiple levels, and would likely elicit synergistic reaction if administered in combination. The present study demonstrates that α-CAP-specific IgY can potentially be used as passive immunotherapeutic for preventing or reducing C. jejuni colonization in chickens. Further in vitro studies will be required to determine the optimal titer of the antibodies for use as passive immunotherapeutic and to demonstrate the in vivo effectiveness of these antibodies.
Footnotes
Acknowledgments
We gratefully acknowledge the technical assistance of Carol Casavant, Jennifer Santos, Kim Lam Chiok, and Lisa Orfe. This project was funded in part with federal funds from the USDA-NIFA PAR-10-276 (2011-67015-30772) and by the Agricultural Animal Health Program, College of Veterinary Medicine, Washington State University.
Disclosure Statement
No competing financial interests exist.