
Abstract
The aim of the present study was to evaluate the anti-Vibrio activity of propolis collected from three different areas of the Sonoran Desert in northwestern, Mexico [Pueblo de Alamos (PAP), Ures (UP), and Caborca (CP)]. The anti-Vibrio spp. activity of Sonoran propolis was determined by the broth microdilution method. UP propolis showed the highest antibacterial activity [minimal inhibitory concentration (MIC50)<50 μg mL−1] against Vibrio spp. (UP>CP>PAP). UP propolis significantly inhibited the growth of Vibrio cholerae O1 serotype Inaba (MIC50<50 μg mL−1), V. cholerae non-O1 (MIC50<50 μg mL−1), V. vulnificus (MIC50<50 μg mL−1), and V. cholerae O1 serotype Ogawa (MIC50 100 μg mL−1), in a concentration-dependent manner. The UP propolis constituents, galangin and caffeic acid phenethyl ester (CAPE), exhibited a potent growth inhibitory activity (MIC50 0.05–0.1 mmol l−1) against V. cholerae strains (non-O1 and serotype Ogawa). The strong anti-Vibrio activity of Sonoran propolis and some of its chemical constituents (galangin and CAPE) support further studies on the clinical applications of this natural bee product against different Vibrio spp., mainly V. cholerae.
Introduction
V
Vibrios may be classified into two groups: “cholera vibrios” and “noncholera vibrios.” The former includes isolates belonging to O1 and O139 of V. cholerae; the latter consists of serogroups other than O1 and O139 and isolates belonging to other Vibrio spp. Cholera vibrios are responsible for cholera, which is a potentially epidemic and severe life-threatening secretory diarrhea characterized by numerous and voluminous watery evacuations, often accompanied by vomiting, and resulting in hypovolemic shock and acidosis (Baron, 1996; Tantillo et al., 2004). Although rehydration is the most important intervention in cholera, the appropriate use of antimicrobials can reduce disease severity and duration of excretion of V. cholerae (Kariuki and Hart, 2001; Bhattacharya, 2003; Okeke et al., 2007). Different antibiotics are used to treat Vibrio infections (tetracycline, doxicycline, ciprofloxacin, furazolidone, and nalidixic acid). The clinical efficacy of many existing antibiotics is being threatened by the emergence of multidrug-resistant Vibrio strains (Kariuki and Hart, 2001; Okeke et al., 2007; Fischbach and Walsh, 2009). Antibiotic resistance is a serious and increasing health problem worldwide, and search for new sources of antibiotics is a global challenge. Natural products derived from plants, marine organisms, and microorganisms are an important source of new antibacterial agents (Alviano and Alviano, 2009; Sharma et al., 2009; Castillo et al., 2011). Several plant extracts have been shown to possess activity against Gram-positive and Gram-negative, aerobic and anaerobic bacteria, including Vibrio spp. (Domingo and López-Brea, 2003; Cowan, 1999; Garcia et al., 2006; Sharma et al., 2009).
Propolis is a natural resinous hive product collected by honeybees from buds, leaf, and exudates of certain trees and plants. Propolis has been used as a folk medicine in many regions of the world (Ghisalberti, 1979; Burdock, 1998; Bankova et al., 2000; Castaldo and Capasso, 2002; Bankova, 2005). It has been shown to possess a wide spectrum of biological activities such as antibacterial (Popova et al., 2005; Uzel et al., 2005; Velazquez et al., 2007), antioxidant (Choi et al., 2006; Ahn et al., 2007), antiviral (Amoros et al., 1994; Kujumgiev et al., 1999), fungicidal (Quiroga et al., 2006; Silici and Koc, 2006), anti-inflammatory (Khayyal et al., 1993), and anticancer activities (Akao et al., 2003; Scifo et al., 2004). The chemical composition of propolis and its biological properties depend on the type of vegetation present in the collection area. Because of the geographical differences, propolis samples from Europe, South America, and Asia vary in chemical composition and activities (Hegazi et al., 2000; Bankova et al., 2002; Usia et al., 2002; Salomao et al., 2004; Isla et al., 2005; Da Silva et al., 2006; Hernandez et al., 2007; Velazquez et al., 2007). Recently, we have described the chemical composition and antibacterial activities of propolis collected from three different arid and semiarid areas of the Sonoran Desert in northwestern Mexico [Caborca (CP), Pueblo de Alamos (PAP), and Ures (UP)] (Hernandez et al., 2007). Sonoran propolis showed strong antibacterial activity against the Gram-positive bacteria Staphylococcus aureus and Listeria monocytogenes. None of the Sonoran propolis tested had significant antibacterial activity against the Gram-negative bacterium Escherichia coli or Pseudomonas aeruginosa (Velazquez et al., 2007).
In the present study, we evaluated the anti-Vibrio activity of propolis collected from three different regions of Sonora, Mexico (CP, PAP, and UP). Study of some of its chemical constituents against this pathogen was another goal of this work.
Materials and Methods
Reagents
Authentic standards of flavonoids—rutin, naringenin, hesperetin, chrysin, and acacetin—were purchased from INDOFINE Chemical Co. (Hillsborough, NJ). Galangin and gentamicin were purchased from Sigma Chemical (St. Louis, MO). Pinocembrin and pinobanksin 3-acetate were isolated from Ures, Sonora, propolis in our laboratory (Hernández et al., 2007). Caffeic acid phenethyl ester (CAPE) was synthesized by chemical reaction of caffeic acid and phenethyl alcohol according to the procedure of Grunberger et al. (1988).
Propolis
Propolis was collected from three different areas of Sonora, Mexico: PAP, UP, and CP. Propolis was collected from November 2003 to March 2004 by professional beekeepers. The hives were located in El Coyote (29° 27’ 04’’ N; 110° 22’ 00’’ W, province of Ures), Los Mesquites (29° 07’ 129’’ N; 110° 07’ 760’’ W, province of Pueblo de Alamos), and Caborca (30o 46’ 844’’ N; 112o 05’ 441’’ W), altitude 413, 652, and 292 m above sea level, respectively. Once collected, propolis was stored at −80°C until used.
Preparation of methanol extracts of propolis (MEP)
MEP were prepared as described previously (Hernandez et al., 2007). Briefly, crude propolis material (100 g) was cut into small pieces and extracted three times with absolute methanol (3×0.4 L) at room temperature for 24 h. After filtration through a filter paper (Whatman grade no. 4), the filtrates were combined and the solvent evaporated under reduced pressure to give dried propolis. The dried propolis was redissolved in absolute methanol and filtered through a nylon membrane (type NYL, 0.2 μm; Whatman, Maidstone, Kent, UK), and the solvent was evaporated in vacuum to give a MEP. The MEP was kept at −80°C in the dark until use.
Bacterial strains
Bacterial strains used in this study were as follows: V. alginolyticus (American Type Culture Collection [ATCC] 17749), V. parahaemolyticus (ATCC 17802), V. vulnificus (ATCC 27562), V. fluvialis (PAHO/WHO), V. cholera O1 serotype Inaba (National Institute for Diagnostics and Epidemiological Reference [InDRE])–Mexico), V. cholera O1 serotype Ogawa (InDRE-México), and V. cholera non-O1 (InDRE-Mexico). Bacterial cultures were maintained in thiosulfate citrate bile salts sucrose (TCBS) broth. Before testing, all bacterial strains were maintained frozen at −70°C in 10% glycerol broth. Identification of Vibrio spp. was carried out on the basis of colony morphology, Gram staining, and standard biochemical tests (oxidase, mobility, T1N0 T1N3, T1N6, T1N8, T1N10, arginine, lysine, ornithine, citrate, hydrogen sulfide [H2S], urea, indol, Voges Proskauer test, gelatinase, and acid production from
Antibacterial assay
In vitro antibacterial studies were carried out by the broth microdilution method as described previously (Velazquez et al., 2007), with some modifications. Briefly, 15 μL (1.5×106 CFU) of a suspension of a logarithmic phase bacterial culture (108 CFU mL−1), the turbidity of this bacterial suspension matching the turbidity of a 0.5 McFarland standard (Velazquez et al., 2007), were inoculated into each well of a flat 96-well microplate (Costar, Corning, NY), containing 200 μL of different concentrations of propolis extracts (50–400 μg mL−1) or its constituents (0.8, 0.4, 0.2, and 0.1 mM) in trypticase soy broth supplemented with 2% NaCl. Propolis (MEP) and its constituents were first dissolved in dimethyl sulfoxide (DMSO) and subsequently diluted in sterile broth. Propolis constituents were tested at different concentrations. Additionally, each antibacterial test included wells containing the culture media plus DMSO (0.2–0.8%), in order to obtain a control of the solvent's antibacterial effect. Gentamicin (12 μg mL−1) was used as positive control of bacterial growth inhibition. Bacterial cultures were incubated at 36°C for 48 h. Plates were read at 620 nm in an enzyme-linked immunoassay (ELISA) microplate reader (Benchmark Microplate Reader; Bio-Rad, Hercules, CA) at different incubation times (6, 12, 24, and 48 h). The optical density (OD620nm) was corrected by subtracting the OD620nm from wells with propolis alone in sterile broth. The minimal inhibitory concentration was defined as the lowest propolis concentrations that inhibited at least 50% (MIC50) or 90% (MIC90) of the bacterial growth after incubation at 36°C for 24 h. MICs were determined using the following criteria (Baizman et al., 2000; Velazquez et al., 2007):
MIC50: (OD620nm untreated bacteria−OD620nm test concentration)/(OD620nm untreated bacteria)×100≥50%
MIC90: (OD620nm untreated bacteria−OD620nm test concentration)/(OD620nm untreated bacteria)×100≥90%
Statistical analysis
Data were analyzed using analysis of variance with Tukey-Kramer and Duncan's multiple comparison tests (Number Cruncher Statistical Software 2000; NCSS, Kaysville, UT).
Results
Antibacterial activity of Sonoran propolis against Vibrio spp.
UP propolis significantly inhibited growth of V. cholerae O1 serotype Inaba (MIC50<50 μg mL−1, MIC90 200 μg mL−1), V. cholerae non-O1 (MIC50<50 μg mL−1, MIC90 200 μg mL−1), V. vulnificus (MIC50<50 μg mL−1, MIC90 200 μg mL−1), and V. cholerae serotype Ogawa (MIC50<100 μg mL−1, MIC90>400 μg mL−1), in a concentration-dependent manner (Fig. 1, Table 1). CP and PAP propolis showed a high antibacterial activity against V. cholerae non-O1 (MIC50<50 μg mL−1, MIC90 400 μg mL−1) and V. cholerae O1 serotype Inaba (MIC50<50 μg mL−1, MIC90>400 μg mL−1). V. alginolyticus, V. fluvialis, and V. parahaemolyticus were the least susceptible to Sonoran propolis (Fig. 1, Table 1). The highest concentration of DMSO used (0.8% v/v) in the antibacterial assays did not interfere with the bacterial growth. The antibiotic gentamicin (12 μg mL−1) was used as a positive control that completely inhibited bacterial growth (Fig. 1).
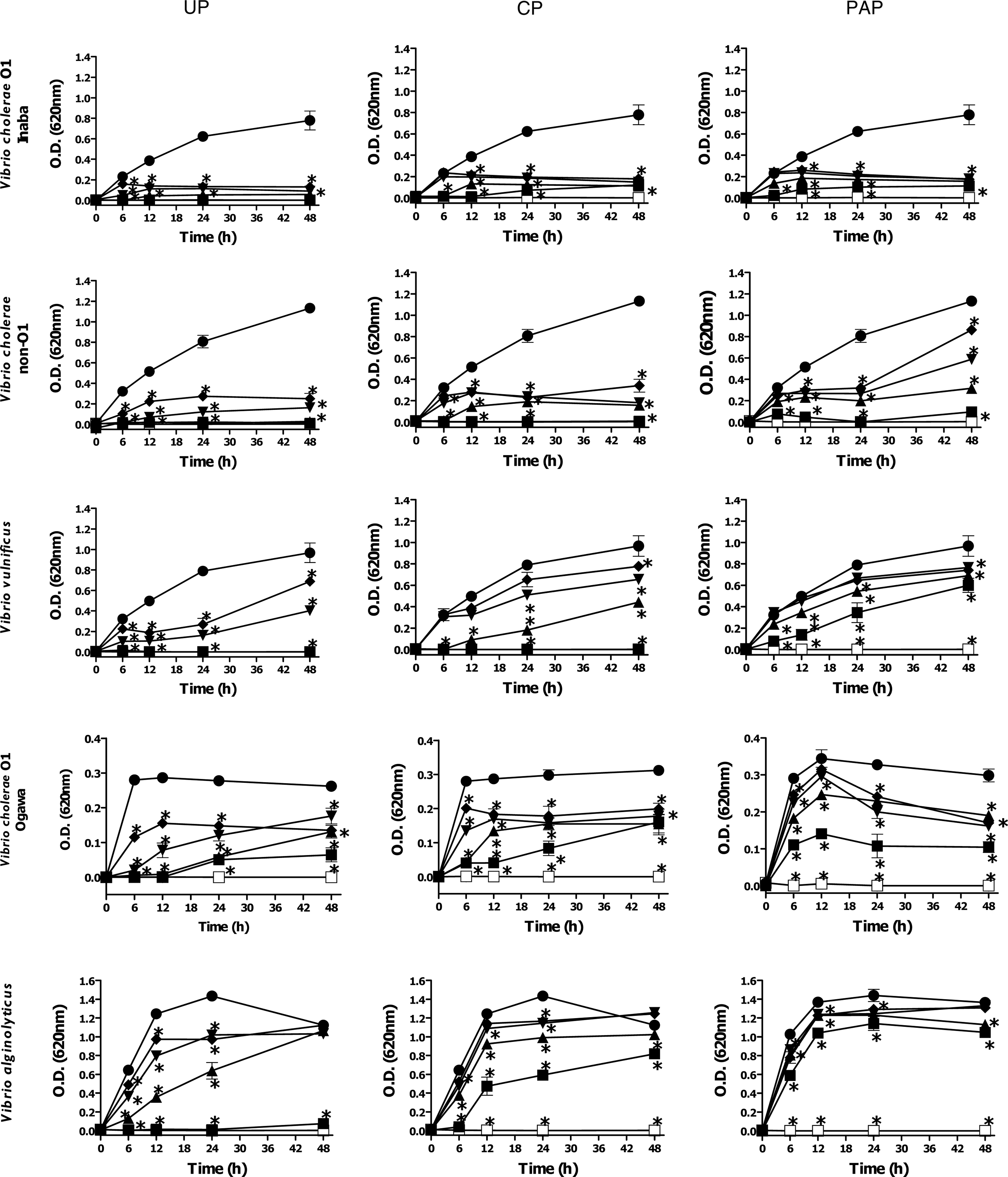
Antibacterial activity of Sonoran propolis Ures (UP), Caborca (CP), and Pueblo de Alamos (PAP) against Vibrio spp. Bacterial cell cultures were treated with different doses of propolis samples during 48 h. Antibacterial effect of propolis was determined by the broth microdilution method as in the text. ■, 400 μg mL−1; ▲, 200 μg mL−1; ▼, 100 μg mL−1; ♦, 50 μg mL−1; ●, 0.0 μg mL−1; □, gentamicin. Control bacterial cultures were incubated with dimethyl sulfoxide (0.2–0.8%). Gentamicin (12 μg mL−1) was used as positive control of bacterial growth inhibition. The results shown are representative of at least two independent experiments. All values represent mean of triplicate determinations±SD. Significant differences (p<0.05) from control cell cultures are marked with an asterisk.
Concentration in μg mL−1.
Anti–Vibrio cholerae activity of Sonoran propolis constituents
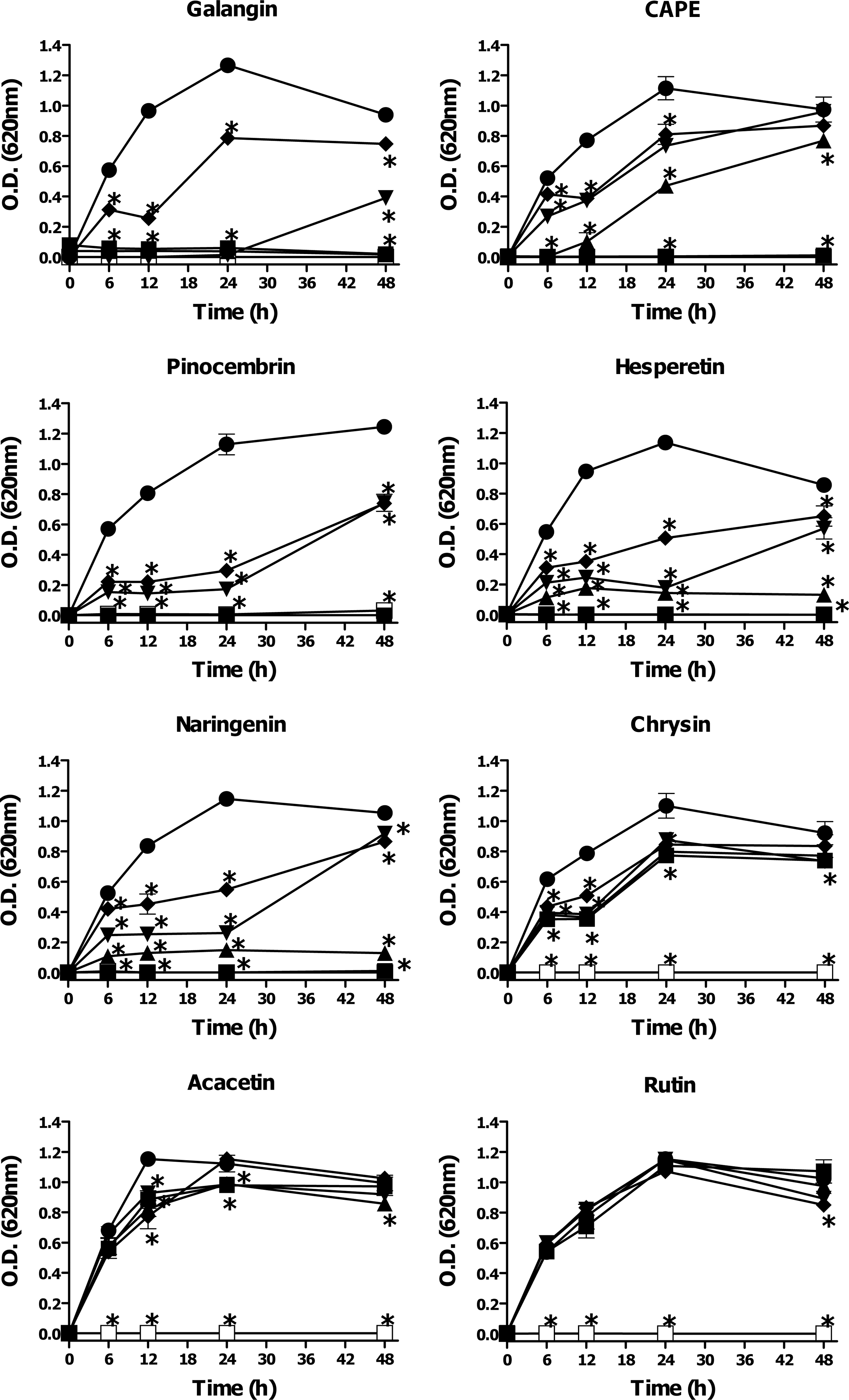
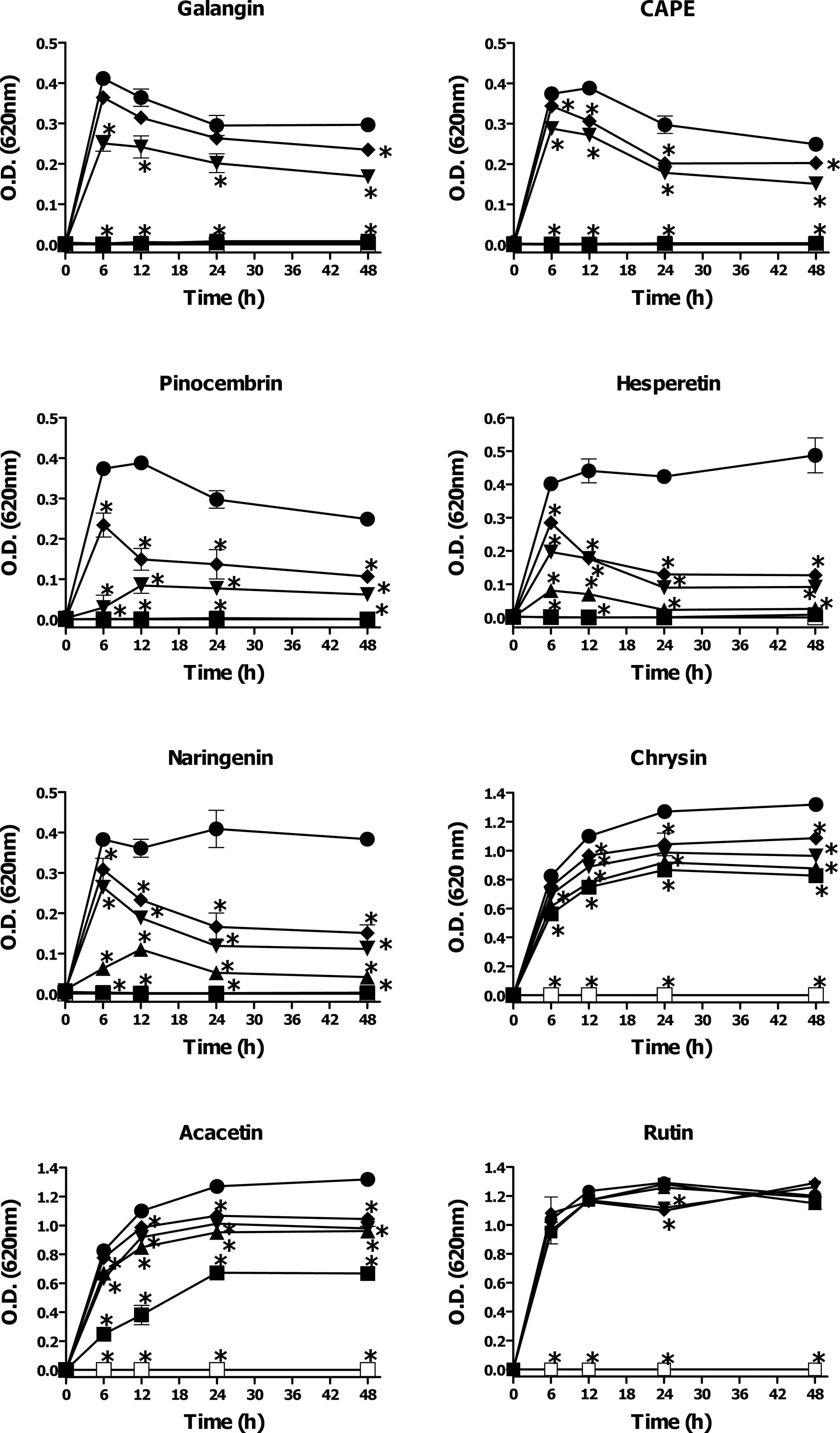
In an earlier study by our group, the major chemical constituents of Sonoran propolis (UP, CP, and PAP) were identified by high-performance liquid chromatography–tandem mass spectrometry (HPLC-MS) as pinocembrin, pinobanksin 3-acetate, chrysin, naringenin, CAPE, acacetin, galangin, pinobanksin, xanthomicrol, 3’-demethoxysudachitin, rutin, and hesperetin (Velazquez et al., 2007). We evaluated the antimicrobial activity of some of the main constituents of Sonoran propolis against V. cholerae O1 serotype Inaba and V. cholerae non-O1. The propolis constituents galangin, CAPE, pinocembrin, hesperetin, and naringenin showed significant inhibitory activity on the growth of both strains of V. cholerae tested. Galangin (MIC90 0.05–0.1 mmol L−1) and CAPE (MIC90 0.2 mmol L−1) exhibited the highest inhibitory effect on the bacterial growth (Figs. 2 and 3, Table 2). Pinocembrin (MIC90 0.4 mmol L−1), hesperetin (MIC90 0.4–0.8 mmol L−1), and naringenin (MIC90 0.8 mmol L−1) had moderate anti-Vibrio activity. Chrysin (MIC90>0.8 mmol L−1) and acacetin (MIC90>0.08 mmol L−1) showed very low inhibitory activity against V. cholerae strains tested. Rutin did not show any significant effect on the growth of V. cholerae O1 serotype Inaba or V. cholerae non-O1 at any of the tested concentrations (0.1–0.8 mmol L−1).
Antibacterial activity of some of the main constituents of Sonoran propolis against Vibrio cholerae non-O1. Bacterial cell cultures were treated with different doses of propolis constituents during 48 h. Antibacterial effect was determined by the broth microdilution method as described in the text. Compounds were evaluated at 0–0.8 mM (▪, 0.8 mM; ▴, 0.4 mM; ▾, 0.2 mM; ♦, 0.1 mM; •, 0.0 mM; □, gentamicin), except for Galangin and CAPE, 0–0.2 mM (▪, 0.2 mM; ▴, 0.1 mM; ▾, 0.05 mM; ♦, 0.025 mM; •, 0.0 mM; □, gentamicin). Control bacterial cultures were incubated with dimethyl sulfoxide (0.2–0.8%). Gentamicin (12 μg mL−1) was used as positive control of bacterial growth inhibition. The results shown are representative of at least two independent experiments. All values represent mean of triplicate determinations±SD. Significant differences (p<0.05) from control cell cultures are marked with an asterisk.
Antibacterial activity of some of the main constituents of Sonoran propolis against Vibrio cholerae O1 Inaba. Bacterial cell cultures were treated with different doses of propolis constituents during 48 h. Antibacterial effect was determined by the broth microdilution method as described in the text. Compounds were evaluated at 0–0.8 mM (▪, 0.8 mM; ▴, 0.4 mM; ▾, 0.2 mM; ♦, 0.1 mM; •, 0.0 mM; □, gentamicin), except for Galangin and CAPE, 0–0.2 mM (▪, 0.2 mM; ▴, 0.1 mM; ▾, 0.05 mM; ♦, 0.025 mM; •, 0.0 mM; □, gentamicin). Control bacterial cultures were incubated with dimethyl sulfoxide (0.2–0.8%). Gentamicin (12 μg mL−1) was used as positive control of bacterial growth inhibition. The results shown are representative of at least two independent experiments. All values represent mean of triplicate determinations±SD. Significant differences (p<0.05) from control cell cultures are marked with an asterisk.
Concentration in mM.
Discussion
In the present study, we evaluated the anti-Vibrio activity of Sonoran propolis collected from three different arid and semi-arid regions of Sonora, Mexico: CP, UP, and PAP. Sonoran propolis had a strong growth-inhibitory activity against several Vibrio spp. This observation is noteworthy because a large number of reports showed that propolis from different geographic areas possesses a weak activity against Gram-negative bacteria (Kujumgiev et al., 1999; Pepeljnjak and Kosalec 2004; Uzel et al., 2005). In addition, we have previously reported that Sonoran propolis did not have antibacterial activity against the Gram-negative bacteria E. coli and P. aeruginosa (Velazquez et al., 2007).
Propolis from Ures, Sonora, showed the strongest growth-inhibitory activity against Vibrio spp. (UP>CP>PAP). The differences observed in the anti-Vibrio activity of the propolis extracts analyzed could be due to variations (qualitative and quantitative) in the chemical composition of the propolis samples (Simoes-Ambrosio et al., 2010). In the present study, we evaluated the antibacterial activity of the major identified compounds of Sonoran propolis against V. cholerae O1 serotype Inaba and V. cholerae non-O1. Galangin and CAPE exhibited a potent inhibitory activity (MIC90 0.05–0.1 mmol L−1) against both V. cholerae strains tested. CAPE is one of the most active compounds of propolis, and it has been mostly detected in propolis from temperate regions (Bankova et al., 2000; Bankova et al., 2002). It has been shown that CAPE has antioxidant, anti-inflammatory, anticarcinogenic, antiviral, immunomodulatory, and other biological properties (Fesen et al., 1994; Su et al., 1994; Natarajan et al., 1996; Michaluart et al., 1999; Okutan et al., 2005; Xiang et al., 2006). In an earlier study, we have shown that CAPE possesses a potent inhibitory activity against S. aureus growth. CAPE was only present in the UP propolis (Hernandez et al., 2007); therefore, CAPE could contribute significantly to the strong anti-Vibrio activity shown by UP. Galangin also showed a high growth-inhibitory activity against V. cholerae, and it was present in the three propolis samples tested. There have been several reports about the antibacterial activity of galagin, mainly against S. aureus. To our knowledge, this is the first study showing that CAPE and galangin possess a potent growth inhibitory activity against V. cholerae. The propolis compounds pinocembrin, hesperetin, and naringenin exhibited a significant effect on the growth of V. cholerae. Pinocembrin is one of the most abundant constituents of Sonoran propolis. This flavonoid is a typical chemical constituent of poplar-type propolis (i.e., propolis from Europe and non-tropic regions of Asia, and it possesses antibacterial action against methicillin-resistant S. aureus (Pepeljnjak and Kosalec, 2004). It has been shown that naringenin has a moderate growth inhibitory activity against V. harveyi (Ulanowska et al., 2006). The inhibitory activity of Sonoran propolis against Vibrio spp. could be due to a synergistic action of some of its constituents (Blonska et al., 2004), resulting in a strong anti-Vibrio activity. Additionally, some of the unidentified compounds present in propolis samples could be responsible for such antibacterial effects. Additional studies need to be carried out to define and characterize at the chemical and biochemical level the inhibitory activity of propolis against Vibrio spp.
It is difficult to draw structure-activity relationships from this limited number of flavonoid compounds, but it may be noted that the hydroxyl group on carbon 3 of galangin has a significant effect on its growth-inhibitory activity, compared to that of chrysin. The double bond at carbon 2–3 of chrysin decreases its inhibitory activity as compared to that of pinocembrine. The hydroxyl group at carbon 4’ on the B ring of naringenin decreases slightly its anti-V. cholerae activity, compared to that of pinocembrin. Further studies need to be conducted to elucidate the structure–activity relationship of the identified propolis compounds.
Conclusion
The results obtained indicate that Sonoran propolis and some of its chemical constituents possess potent in vitro anti-Vibrio spp. activity. Additional studies are needed to explore the possible use of this natural product and its constituents to prevent and/or control the water- and foodborne diseases caused by V. cholerae. Complete chemical characterization of Sonoran propolis should provide a better understanding of the biological activity of this product.
Footnotes
Acknowledgments
We thank professional beekeepers Humberto Saavedra, Gilberto Valenzuela, and Jose Puentes for propolis collection.
Disclosure Statement
No competing financial interests exist.