
Abstract
In this study, we investigated the molecular mechanisms involved in erythromycin resistance in the first resistant Campylobacter strains isolated from chicken meat in Poland, and analyzed their genetic relatedness. A total of 297 samples of raw chicken meat and giblets from retail trade in the Warsaw area collected between 2006 and 2009 were examined. Among 211 Campylobacter strains (52 C. jejuni and 159 C. coli), 10 C. coli isolates (4.7%) were resistant to erythromycin. All the C. jejuni strains were susceptible. Among the high-level macrolide-resistant isolates, two different point mutations within the domain V of the 23S rRNA gene were observed. Eight of the strains had adenine→guanine transitions at position 2075, two other isolates at position 2074. Sequence analysis of ribosomal proteins L4 (rplD) and L22 (rplV) indicated that ribosomal protein modifications did not contribute to macrolide resistance. A mutation in the inverted repeat in the cmeR and cmeABC intergenic region was found in a single resistant strain. The genetic relatedness of Campylobacter isolates showed that two resistant strains obtained from the same production plant in a 2-month interval were genetically identical. The risk of transmission of resistant strains via the food chain highlights the need for constant monitoring of resistance in Campylobacter isolates of human and animal hosts.
Introduction
Campylobacteriosis is generally a self-limiting disease. Antimicrobial treatment is necessary for systemic Campylobacter infections, infections in immune-suppressed patients, and severe or long-lasting infections. Macrolides are considered the drugs of choice for treating Campylobacter infections (Allos, 2001). There is, however, serious concern associated with the increasing frequency of isolation of resistant Campylobacter spp. from various sources such as humans, animals, and food (Kurincic et al., 2007). Consequently, European Union countries are required as a result of the decision of Committee 2007/516/WC to monitor the antimicrobial resistance of Campylobacter spp. in broiler flocks and the prevalence of Campylobacter spp. in broiler carcasses.
Two main mechanisms can be involved in macrolide resistance in Campylobacter (i.e., ribosomal target modifications and active efflux) (Gibreel and Taylor, 2006). Ribosomal target modifications can result from point mutations in the peptidyl-encoding region in the domain V of the 23S rRNA gene, or mutation in the ribosomal proteins L4 and L22. High-level resistance to erythromycin is associated with point mutations at positions 2075 or 2074 of the 23S rRNA gene (corresponding to positions 2058 and 2059 in Escherichia coli) (Gibreel et al., 2005). Modifications in the ribosomal proteins L4 and L22 can possibly be associated with low-level resistance; however, to date there is no evidence of their contribution to high-level resistance (Cagliero et al., 2006; Caldwell et al., 2008). Macrolide resistance can be also mediated by the CmeABC efflux pump, which protects Campylobacter against antimicrobials and other toxic compounds such as bile salts, detergents and dyes (Lin et al., 2002). The expression level of the CmeABC influences the level of macrolide resistance and may act synergistically with other resistance mechanisms (Gibreel et al., 2005; Cagliero et al., 2006).
The aim of this work was to analyze the molecular mechanisms involved in macrolide resistance in C. jejuni and C. coli strains isolated from raw chicken meat and giblets in Poland in the years 2006–2009, with focus on region V of the 23S rRNA gene, the rplD and rplV genes, and the regulatory region of the cmeABC operon (CmeABC efflux pump), after first establishing the antibiotic susceptibilities of the strains.
Materials and Methods
Sample collection
A total of 297 food samples were obtained from large retail markets and smaller units in the Warsaw area during 2006–2009. Two hundred forty samples of raw chicken meat (wings, legs, carcass frames, fillets, ground meat) and 57 samples of giblets (livers, hearts, gizzards) were sampled. Only fresh, previously nonfrozen samples were taken. After collection, the samples were transported to the laboratory in a portable cooler at 2–8°C and immediately subjected to microbiological analysis.
Isolation and species identification
A 10-g portion of each sample of chicken meat was analyzed in accordance with standard EN ISO 10272-1:2006 procedure (qualitative analysis). After initial bacterial characterization by phase-contrast microscopy, catalase and oxidase production, and growth at 25°C and 42°C, suspected colonies were identified as C. jejuni or C. coli using the polymerase chain reaction (PCR) method with species-specific primers, as described by On and Jordan (2003) (Table 1).
Prior to analysis all Campylobacter isolates were maintained at −70°C in Brucella broth containing 20% glycerol.
Antibiotic susceptibility testing
Initial screening of susceptibility to erythromycin of 211 Campylobacter isolates was performed on Mueller-Hinton agar plates containing 5% sheep blood (PB5007A; Oxoid, Basingstoke, Hampshire, UK) using discs containing erythromycin (15 μg) (Becton Dickinson, Sparks, MD). Briefly, colonies from 48-h cultures were suspended in Mueller–Hinton broth to a turbidity of 0.5 McFarland units. This suspension was plated on a Mueller–Hinton agar plate with 5% sheep blood, and a disc was put on each plate after the surface of the plate had dried. The plates were incubated under microaerophilic conditions (CampyGen, Oxoid) at 37°C for 48 h. No inhibition zone and diameters of ≥15 mm around the erythromycin disk were suggested to be resistant and susceptible breakpoints, respectively, of Campylobacter spp. strains (Gaudreau et al., 2008).
Susceptibility to erythromycin, tetracycline, and ciprofloxacin isolates considered resistant to erythromycin by the disk-diffusion method was determined by the Etest method (AB Biodisk, Solna, Sweden). To ensure reproducibility, minimum inhibitory concentration (MIC) determinations were repeated at least twice. The MIC breakpoints used were the following: >8 for erythromycin, >2 for tetracycline, and >.05 μg/mL for ciprofloxacin as recommended by the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2012).
Analysis of the molecular mechanisms of macrolide resistance
Mutations in the 23S rRNA gene were detected by the PCR–restriction fragment length polymorphism method and direct sequencing as described earlier (Vacher et al., 2003). Modifications in L4 and L22 ribosomal proteins of Campylobacter were analyzed as described by Corcoran et al. (2006). DNA sequences of resistant and susceptible strains were compared with the sequence of C. coli JV20 genome (GeneBank accession number NZ_ AEER01000024).
The cmeR gene (636 bp), intergenic region between cmeR and cmeA (107 bp), and the first part of cmeA (82 bp) were amplified with a pair of new designed primers: CmecoliF1 (5′ATGAATCCAAATAAAACACC 3′) and CmecoliR2 (5′ GAGGTGCTTCTTCTTTGCTAC 3′) on the basis of previously sequenced bacteria (GeneBank accession number FJ797670-3) (Guo et al., 2010). PCR consisted of 30 cycles of 94°C for 30 s, 45°C for 30 s, and 72°C for 2.30 min.
All PCR products were purified using QIAquick PCR Purification KIT (Qiagen, Germany) following the manufacturer's instructions. Sequencing was performed using DYEnamic ET Terminator Cycle Sequencing Kit (GE Healthcare, Chalfont St. Giles, UK) on an Applied Biosystems 3730 automated sequencer (Applied Biosystems, Perkin-Elmer, Foster City, CA).
The sequences of all primers used for molecular analysis are listed in Table 1.
Analysis of genetic similarity of macrolide-resistant strains
The genetic relatedness of Campylobacter isolates resistant to erythromycin was analyzed by the pulsed-field gel electrophoresis (PFGE) method as described by Ribot et al. (2001) using a CHEF-DRIII system (Bio-Rad, Hercules, CA) for electrophoresis. The DNA fragments, after digestion with the SmaI (Thermo Scientific Fermentas, Vilnius, Lithuania), were separated for 18 h with a pulse time ramped from 6.8 s to 38 s at a temperature of 14°C. The PFGE patterns were analyzed using Molecular Analyst Software Fingerprinting (Bio-Rad). The macrorestriction patterns were normalized by interpolation to the nearest referent line. Dice's similarity coefficients were calculated to generate matrix and dendrogram using the unweighted pair group method with arithmetic mean method. Band position tolerance was of 2.0% and optimization of 0.5%. Isolates were considered as belonging to the same cluster when the similarity coefficient was >90%.
Results
Antimicrobial resistance
A total of 211 Campylobacter isolates from chicken samples were collected during the study period (159 C. coli and 52 C. jejuni). Ten of the tested Campylobacter strains (4.7%) showed phenotypic resistance to erythromycin. All of them were identified as C. coli.
The erythromycin MICs of all resistant isolates were ≥256 μg/mL (Table 2). The MICs against four erythromycin-susceptible isolates C. coli and reference strain C. coli NCTC 11366 ranged from 0.5 to 4 μg/mL.
Isolates were considered resistant to tetracycline (TET), ciprofloxacin (CIP), and erythromycin (ERY) with MICs>2, >0.5, and >8 μg/mL, respectively (EUCAST, 2012).
The position of the 23S rRNA gene mutation.
The position of amino acids changes. DNA sequences of rplD and rplV genes coding L4 and L22 ribosomal proteins, respectively, were compared with the sequence of C. coli JV20 genome (GeneBank accession number NZ_ AEER01000024).
Erythromycin-susceptible C. coli strains used for comparison.
In addition to macrolide resistance, eight C. coli were also resistant to tetracycline and ciprofloxacin. Two strains—168/06 and 251/08—were resistant to ciprofloxacin (MIC>32 and 8, respectively) but were susceptible to tetracycline (MIC 0.5 and 2).
Analysis of 23S rRNA sequences
PCR–restriction fragment length polymorphism and direct sequencing were used for the detection of 23S rRNA mutations in 10 C. coli strains phenotypically resistant to erythromycin and in five susceptible ones (Table 2).
Among the resistant strains, two different point mutations within the domain V of the 23S rRNA gene were observed. Mutations in highly resistant strains were identified at position 2074 and 2075. Eight of the 10 C. coli isolates exhibited the adenine-to-guanine transition at position 2075 (A2059G, E. coli numbering) while two had A2074G transition. No mutation was identified in this region in any of the susceptible isolates.
Based on sequence results, it appeared that the A2074G and A2075G mutations were present in all three copies of the 23S rRNA gene because the sequence chromatograms showed only a high peak for guanine in that position.
Analysis of the L4 and L22 ribosomal protein
All of 10 macrolide-resistant strains and five susceptible ones were examined regarding alterations in the genes encoding ribosomal proteins L4 and L22. None of resistant strains harbored any modification in L4, while substitutions in L22 were identified: A103V (three isolates), T109A (four isolates), and V129A (two isolates). It was interesting to note that T109A and A103V mutations were also found in one susceptible isolate (Table 2).
Analysis of the regulatory region of cmeABC
To investigate the contribution of efflux pump activity to high-level macrolide resistance, the changes in the cmeR gene and cmeR-cmeA intergenic region were analyzed. Sequence analysis indicated that the cmeR gene in resistant strains was identical to the one in susceptible strains.
The 107 nucleotides long cmeR-cmeA intergenic region was located from −50 to −35 upstream of the cmeA gene (consensus sequence:
PFGE analysis of macrolide-resistant strains
Resistant C. coli isolates were genotyped using the PFGE method to analyze their genetic relatedness. Molecular analysis showed nine different macrorestriction patterns (Fig. 1). Two strains, 273/09 and 292/09, had identical patterns. Two other isolates (251/08 and 357/09) showed high (>90%) genetic similarity and have been classified to the same group.
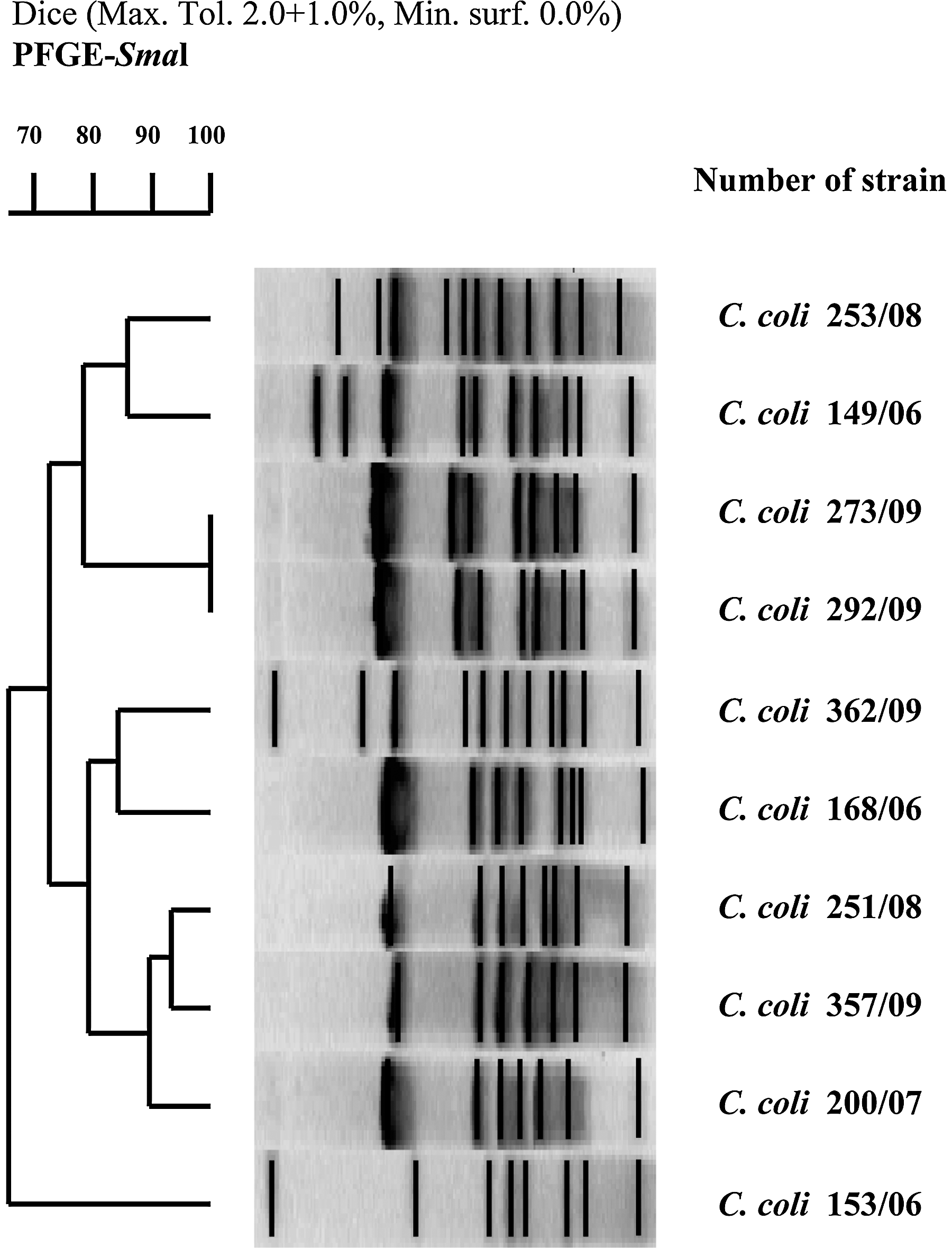
Pulsed-field gel electrophoresis (PFGE) profiles of 10 Campylobacter coli isolates resistant to macrolides.
Discussion
The prevalence of macrolide resistance in Campylobacter strains varies depending on geographical region. The rate of erythromycin resistance in C. jejuni isolated from broiler meat in reporting European countries in the period 2006–2009 was 1%, 3%, 6%, and 3% respectively. While resistance levels to erythromycin in C. coli were 5% in 2006, 6% in 2007, and 13% in 2009 (EFSA, 2010a; EFSA, 2010b; EFSA, 2011).
The previous study conducted in Poland in 2003–2005 did not find macrolide resistance in C. jejuni and C. coli isolates of chicken origin (Rożynek et al., 2008). In this study, the rate of resistance to erythromycin was low overall (4.7%) and similar to that reported in poultry isolates in other European countries. C. coli strains were more resistant to macrolides (6.3%; 10/159) than C. jejuni (0%; 0/52).
Currently, it is considered that the use of macrolides in food-producing animals at therapeutic or subtherapeutic concentrations is a major risk factor influencing the emergence of erythromycin-resistant Campylobacter (Gibreel and Taylor, 2006; Ladely et al., 2007; Lin et al., 2007).
In this study, the primary mechanism of macrolide resistance was due to a single point mutation in the 23S rRNA as previously reported (Vacher et al., 2003; Payot et al., 2004; Gibreel et al., 2005; Corcoran et al., 2006; Lin et al., 2007; Ladely et al., 2009; Logue et al., 2010; Pérez-Boto et al., 2010). Eight of the 10 C. coli isolates exhibited the A2075G transition, which has been the most common mutation identified among macrolide-resistant Campylobacter isolates (Gibreel and Taylor, 2006). This mutation, as other authors have shown (Gibreel et al., 2005; Pérez-Boto et al., 2010; Lehtopolku et al., 2011), seems to provide some biologically advantageous features. Another macrolide-associated mutation (A2074G transition) was found in only two isolates. The occurrence of mutation A2074G has been described in a few reports (Jensen and Aerostrup, 2001; Niwa et al., 2001; Lin et al., 2007; Caldwell et al., 2008; Logue et al., 2010; Pérez-Boto et al., 2010). Gibreel et al. (2005) have suggested that the reason that A2074G transition occurs at a very low frequency among different resistant isolates is likely the negative effect of the mutation on the growth rate of Campylobacter. However, Lin et al. (2007) have observed no influence of mutation A2074G on the growth of the resistant strains selected in vivo. It is likely that the emergence of a specific point mutation conferring high-level resistance is dependent on the genetic features of a strain, the environment in which the mutation is selected and/or specific macrolide agent used for selection (Lin et al., 2007).
Mutation in the genes encoding the L4 and L22 ribosomal proteins that affect the binding of macrolide antibiotics to the 50S ribosome subunit and thus confer resistance have been identified in some bacteria (Gibreel and Taylor, 2006; Belanger and Shryock, 2007). Some isolates of macrolide-resistant Campylobacter with 23S rRNA mutations have also been shown to contain specific changes in ribosomal protein L4 and L22, but the significance of this finding remains obscure (Gibreel et al., 2005; Corcoran et al., 2006; Hao et al., 2009; Pérez-Boto et al., 2010). On the other hand, some findings have demonstrated that modifications in L4 (G57D/V and G74D) and L22 (insertions at position 86 or 98) might confer macrolide resistance in Campylobacter (Cagliero et al., 2006; Caldwell et al., 2008).
In our study, we have found one amino acid substitution, V121A, in L4 in a macrolide-susceptible isolate. Similarly, a mutation has been previously identified by Corcoran et al. (2006) and Pérez-Boto et al. (2010) in resistant and susceptible strains. Several other nonsynonymous substitutions were identified in L22. The mutations at position 103 and 109 have also been described by others (Corcoran et al., 2006; Pérez-Boto et al., 2010). However, neither of them has been associated with macrolide resistance (Corcoran et al., 2006; Pérez-Boto et al., 2010). All these data together suggest that these particular mutations do not contribute to erythromycin resistance among the collection of C. coli isolated examined.
Antimicrobial resistance in Campylobacter isolates can also result from active drug removal mediated by the CmeABC efflux system (Lin et al., 2002). The expression of this resistance-nodulation-division type of efflux pump is regulated by a transcriptional repressor CmeR, encoded by the cmeR gene. CmeR protein represses the transcription of the cmeABC operon by binding to the promoter region (specifically to the inverted repeat) of the efflux operon (Lin et al., 2005b). Mutations in CmeR or in the inverted repeat have been associated with overproduction of the MDR efflux pump (Lin et al., 2005a). It is generally accepted that CmeABC is linked to low and intermediate resistance to erythromycin, while the 23S rRNA mutation can confer a high level of resistance (Payot et al., 2004; Mamelli et al., 2005). Another study concluded that the 23S rRNA mutation and CmeABC work together in Campylobacter strains to confer high-level macrolide resistance (Cagliero et al., 2005; Gibreel et al., 2007; Lin et al., 2007). In addition, the synergy between the CmeABC efflux pump and mutation in the ribosomal proteins L4 and L22, was also shown to confer macrolide resistance in C. coli and C. jejuni (Cagliero et al., 2006; Caldwell et al., 2008).
In this study, we also analyzed the sequence of the cmeR gene and cmeR-cmeA intergenic region in macrolide-susceptible and resistant isolates. To our knowledge, a single nucleotide deletion between the two half sites of the inverted repeat reduced the level of CmeR binding to the promoter sequence and resulted in overexpression of cmeABC (Lin et al., 2005a). We found the mutation in the IR region of a single resistant strain. Therefore, a misregulation of the operon could be expected in this strain (Lin et al., 2005a). These results indicate that the efflux system may contribute to macrolide resistance in only one C. coli strain investigated in this study.
A recent study conducted in Poland in 2010 showed the possible transmission of resistant Campylobacter strains between humans and the environment (Rożynek et al., 2010). In this study, we also analyzed the genetic relationship of erythromycin-resistant C. coli. Strains having identical patterns were isolated in March and May 2009 from chicken meat from the same producer who distributes his products nationwide. Two other isolates belonging to a second clonal group were isolated from chicken meat from two independent producers. The wide distribution of meat products contaminated with resistant Campylobacter strains may lead to the spread of macrolide resistance, which may have grave public health consequences.
Conclusions
In this study, we examined the mechanisms of macrolide-resistance in erythromycin-resistant Campylobacter strains isolated from chicken meat in Poland. The resistance resulted from an A-G transition at position 2074 and 2075 in the 23S rRNA gene, the main described mechanism among Campylobacter, whereas modifications in the L4 and L22 ribosomal proteins were not involved. We also found a mutation in the cmeR-cmeA intergenic region, which may lead to enhanced tolerance to macrolides.
The risk of the transmission of resistant strains via the food chain stresses the need for the constant monitoring of resistance in Campylobacter human and animal isolates.
Footnotes
Acknowledgments
This work was supported by grant no. 163/06 from the Children's Memorial Health Institute.
Disclosure Statement
No competing financial interests exist.