
Abstract
Strains of the enteric pathogen Vibrio parahaemolyticus harboring the thermostable hemolysin (TDH) encoding gene tdh is known to cause epidemic and pandemic diarrhea. In industrialized countries, this pathogen causes sporadic or outbreaks of diarrheal illness associated with consumption of raw or improperly cooked seafood. This report describes a foodborne outbreak of gastroenteritis caused by V. parahaemolyticus in June 2011 following consumption of food served at a funeral reception held at Habra, North 24 Parganas, West Bengal, India. About 650 people attended the function, of whom 44 had acute watery diarrhea with other clinical symptoms; 35 of them were admitted to the District Hospital for the rehydration treatment. Stool specimens collected from three hospitalized cases were positive for V. parahaemolyticus, of which two strains were identified as an O4:K8 serovar and one was identified as O3:K6 serovar. The O3:K6 strain also possessed the pandemic group-specific toxRS gene target (GS), whereas the O4:K8 strains were negative. All strains were polymerase chain reaction–positive for tdh but were polymerase chain reaction–negative for trh. All of the strains were resistant to ampicillin but were pansensitive to other antimicrobials tested. Pulsed-field gel electrophoresis (PFGE) analysis using NotI showed that the O3:K6 strain was similar to that of a recent clinical strain from Kolkata, but had diverged from other strains during previous years. In contrast, PFGE analysis showed that the O4:K8 strains were closely related but differed from the Kolkata strain.
Introduction
V
Recently, V. parahaemolyticus has received global attention due to the emergence of a new O3:K6 clone, which is the first documented serovar related to a pandemic spread (Matsumoto et al., 2000). The rapid spread of the O3:K6 strains to different countries after its first discovery in Kolkata, India further strengthened the pandemic concept of V. parahaemolyticus infection (Matsumoto et al., 2000). Remarkably, the pandemic strain O3:K6 has seven nucleotide changes in the toxRS operon compared to other serovars (Matsumoto et al., 2000). Based on this strain-specific toxRS polymorphism, a group-specific polymerase chain reaction (GS-PCR) was developed along with the confirmation of tdh, but not the trh gene for the identification of the pandemic O3:K6 strains (Matsumoto et al., 2000). Subsequently, besides O3:K6, other serovars such as O4:K68, O1:K25, O1:K41, O1:K untypeable (UT), O1:K56, O3:K75, O4:K8, O4:K12, O4:KUT, and O5:KUT having the pandemic strain genetic properties have also emerged and have spread to many countries (Chowdhury et al., 2004; Laohaprertthisan et al., 2003; Nair et al., 2007).
Epidemiological studies conducted in Kolkata showed the association of V. parahaemolyticus O3:K6 serovar in a foodborne diarrheal outbreak in 2003 (Sen et al., 2007). In this article, we describe an unusual large foodborne outbreak caused by V. parahaemolyticus in West Bengal, India on June 22, 2011 after the consumption of food served during a funeral ceremony. V. parahaemolyticus strains isolated during the outbreak were characterized serologically and molecularly as well as for antimicrobial susceptibility determination.
Materials and Methods
Background of the outbreak
In June 21, 2011 about 650 people attended a funeral ceremony at Iswarigacha, Habra, North 24 Parganas, West Bengal. Among them, 44 developed typical symptoms of gastroenteritis including watery diarrhea with or without abdominal pain and vomiting after consumption of some nonvegetarian dishes prepared by a caterer. Some of the patients were admitted at the District Hospital and the majority of them were treated at their homes. An investigation coordinated by the West Bengal State Department of Health detected an index case on June 22, 2011 at 4:00 a.m. Of 44 cases, 35 were admitted to the hospital and nine were treated as outpatients. Females were more affected (68.2%) than males. Three rectal swabs and water samples were collected and sent to the laboratory on June 23, 2011. Water samples collected from the tap water supplied from the bore well-water source were tested. However, no food or served water was available for testing. The outbreak was declared over on June 26, 2011.
Stool culture
Nonbloody diarrheal samples were collected as rectal swabs and transported to the laboratory in Cary-Blair medium. The rectal swabs were processed for common enteric pathogens such as pathogenic vibrios, salmonellae, shigellae, campylobacters, and diarrheagenic Escherichia coli using enrichment and selective plating media (WHO, 1987). After enrichment of the rectal swabs in alkaline peptone water (pH 8.6) for 6 h at 37°C, the cultures were plated onto thiosulfate citrate bile salt sucrose agar (Eiken, Tokyo, Japan). Sucrose–nonfermenting green colonies on thiosulfate citrate bile salt sucrose suspected as V. parahaemolyticus were subsequently confirmed according to standard methods as described by WHO (1987) and constitute testing for sucrose, lactose, and glucose fermentation (TSI agar slant), hemolytic, urease, and oxidase activities.
Serotyping
Serotyping of V. parahaemolyticus was done using a commercially available antisera test kit (Denka Seiken, Tokyo, Japan) that has 11 O and 71 K antisera. Following manufacturer's instructions, an aliquot of the cell suspension in normal saline from a Luria agar culture was boiled for 2 h and used for O serotyping. The unboiled cell suspension was used to detect each strain's capsular antigen (K) using the K antisera described in the commercially available serological typing kit (Denka Seiken).
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using the disk diffusion method with commercially available disks such as ampicillin (10 μg), co-trimoxazole (25 μg), ciprofloxacin (5 μg), furazolidone (100 μg), norfloxacin (10 μg), nalidixic acid (30 μg), streptomycin (10 μg), tetracycline (30 μg), and erythromycin (15 μg) (Becton Dickinson, Sparks Glencoe, MD) in accordance with the criteria recommended by Clinical and Laboratory Standards Institute (CLSI, 2011). Since there is no interpretive criterion for V. parahaemolyticus in the CLSI guidelines, breakpoints for Enterobacteriaceae were used to determine antimicrobial susceptibility. Escherichia coli ATCC 25922 was used as a quality control strain.
PCR assays
PCR assays were performed to detect the toxR, tdh, and trh genes by using the primers described previously (Chowdhury et al., 2004). GS-PCR was performed to detect the pandemic strain of V. parahaemolyticus (Chowdhury et al., 2004). Template DNA was prepared using 2 mL growth of the test strains in Luria broth (Difco) supplemented with 3% NaCl. The bacterial cells were washed and resuspended in 200 μL of sterile distilled water. The cells were lysed and the DNA was denatured after boiling the cell suspension for 10 min. The cell supernatants were used as templates in the PCR assays. The PCR amplicons were electrophoresed in a 1% agarose gel stained with ethidium bromide and visualized under ultraviolet light using a gel documentation system (Geldoc 2000, BioRad, Hercules, CA).
Pulsed-field gel electrophoresis (PFGE)
PFGE was performed according to the PulseNet protocol (Parsons et al., 2007) with slight modification (Kam et al., 2008). DNA digestion with restriction endonuclease NotI (25 U/plug; Fermentas, Germany) was performed at 37°C overnight. The digested DNA of Salmonella Braenderup strain H9812 with XbaI was used as a molecular weight marker. The restriction fragments were resolved on a CHEF Mapper system (BioRad) in 0.5X Tris-borate-EDTA buffer (TBE) with 300 μM thiourea and the following running conditions: 6 V/cm for 18 h at 14°C, with 2–40 s switch time and pump speed of 0.7 L/min. Gels were stained with ethidium bromide (25 min), destained in distilled water for 45 min and photographed under ultraviolet light. The PFGE patterns were analyzed using the BioNumerics version 4.0 software (Applied Maths, Sint Martens Latem, Belgium) after normalization of the TIFF images with Salmonella enterica serotype Braenderup size standard. Clustering was performed using the unweighted pair group method (UPGMA) and the Dice correlation coefficient with a position tolerance of 1.5%. The PFGE profiles of O3:K6, O4:K8 from the previous collection were made for clonal comparison.
Results
The outbreak peaked during the morning of June 22, 2011. The last case also occurred on the same day. All of the admitted cases were treated intravenously or with oral rehydration fluids and, depending on severity, some patients were given divided dozes (400 mg each) of norfloxacin. There were no fatalities associated with this outbreak.
Rectal swabs collected from three patients were sent to the laboratory for screening common enteric pathogens. The three rectal swabs collected from the patients were positive for V. parahaemolyticus and no other enteric pathogen was detected. Patients positive for V. parahaemolyticus were adult males aged between 20 and 40 years. The V. parahaemolyticus isolates exhibited hemolytic activity on sheep blood agar but were urease negative.
V. parahaemolyticus strains determined by slide agglutination showed that one was identified as an O3:K6 serovar and the other two strains belonged to the O4:K8 serovar group (Table 1). PCR results for the identification of the pandemic strain and virulence genes are shown in Table 1. The O3:K6 strain was PCR positive for the pandemic GS target gene, confirming its similarity with the pandemic strains, but the serovar O4:K8 strains were negative for this gene. All three V. parahaemolyticus strains harbored the tdh, but not the trh gene (Table 1). Antibiotic susceptibility analysis showed that all the strains were resistant to ampicillin and streptomycin but susceptible to tetracycline, trimethoprim-sulfamethoxazole, ciprofloxacin, norfloxacin, and nalidixic acid.
GS-PCR, group-specific polymerase chain reaction; AM, ampicillin; S, streptomycin.
To determine the relatedness among V. parahaemolyticus O3:K6 (NICED 459) and O4:K8 strains (NICED 458 and NICED 460) isolated in this outbreak, four O3:K6 strains from the Kolkata outbreak that occurred in 2003 (SC 188, SC 189, SC 192, SC193), two strains of O3:K6 (IDH 03525), and O4:K8 (IDH 03062) from diarrheal patients from the Infectious Diseases Hospital (IDH), Kolkata isolated during in 2011 were compared by using PFGE analysis. Cluster analysis showed that the recent outbreak O3:K6 V. parahaemolyticus strain possessed an identical profile with that of the IDH 03525 (O3:K6) strain, but its banding pattern differed from the 2003 outbreak strains (Fig. 1). The current O4:K8 strains, on the other hand, were closely related but distinct from IDH 03062 (O4:K8) isolate (Fig. 1). The O3:K6 and O4:K8 strains formed distinct serovar-related clusters (A and B); hence they are genetically different (Fig. 1).
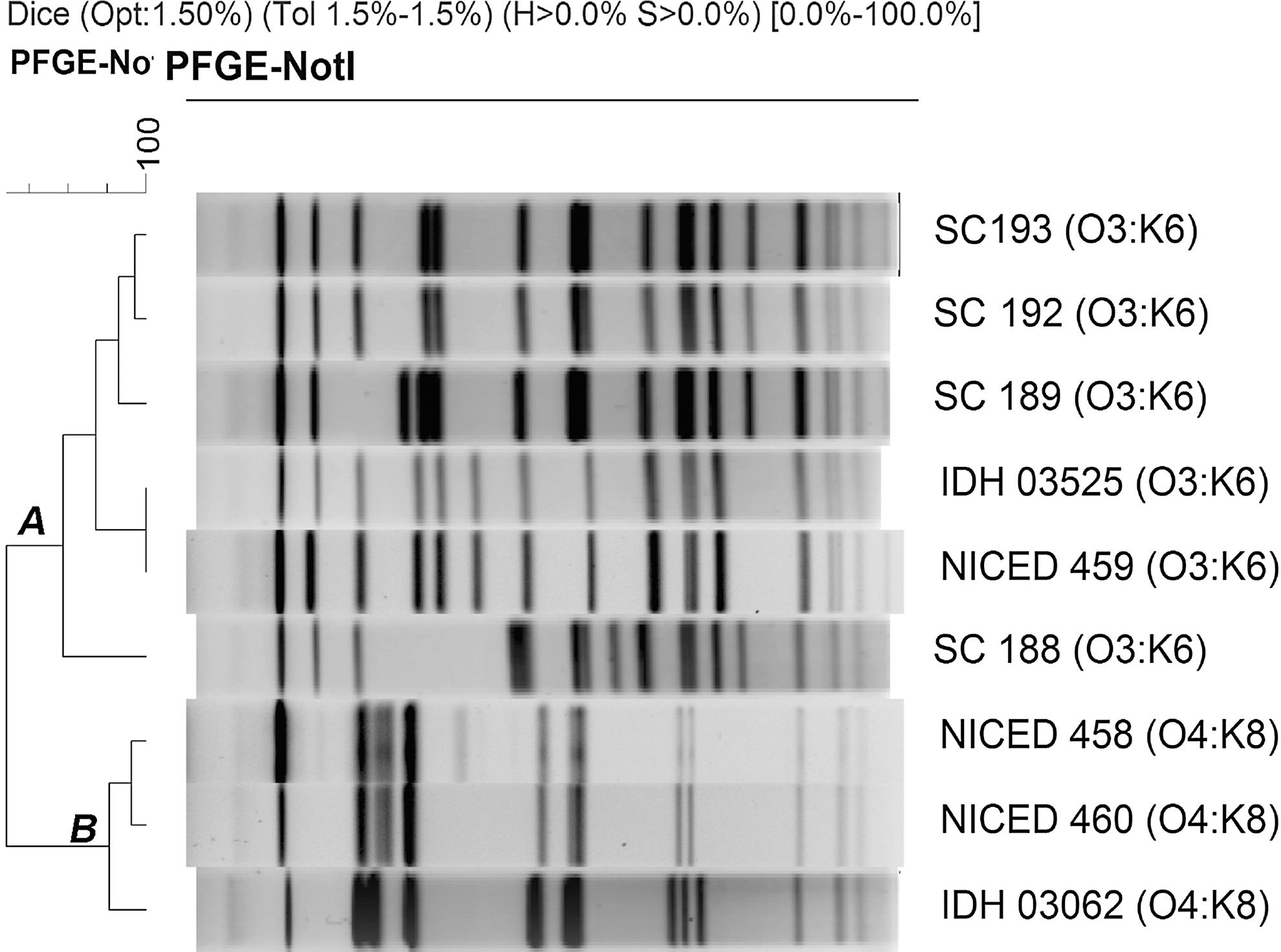
NotI digested chromosomal DNA banding patterns of Vibrio parahaemolyticus O3:K6 and O4:K8 strains. Strain NICED 459 was isolated during the outbreak in 2011 while IDH 03525 was the recent V. parahaemolyticus isolated from Kolkata in 2011. Strains SC188, 189, 192, and 193 were isolated during the outbreak in Kolkata, 2003 (cluster A). Strain NICED 458 and 460 was isolated during the outbreak in 2011, while IDH 03062 was the recent V. parahaemolyticus isolated from Kolkata in 2011 (cluster B). PFGE, pulsed-field gel electrophoresis.
Discussion
The pandemic serovars of V. parahaemolyticus emerged and have become a persistent pathogen associated with diarrhea in many countries. In Kolkata, even though the isolation rate of V. parahaemolyticus is low (∼3%) among the hospitalized diarrheal patients, the pandemic strains persist with frequent changes in the serovars (unpublished data). The pandemic serovar O3:K6 has emerged in Kolkata in 1996 (Matsumoto et al., 2000) and has caused an outbreak in Kolkata during 2003 (Sen et al., 2007). Systematic surveillance conducted in the United States suggests that the burden of diarrhea is increasing due to infection caused mainly by V. parahaemolyticus (Newton et al., 2012). In addition, several other investigations carried out all over the world indicate the association of consumption contaminated seafood with the pandemic strains of V. parahaemolyticus (Nair et al., 2007).
In the present investigation, we identified two serovars of V. parahaemolyticus among three strains isolated in this study: one related to a pandemic O3:K6 strain that was associated with an outbreak in 2011 in Kolkata, and the other two strains (O4:K8) were distinct strain unrelated to previously isolated strains. During 2009, these two serovars were associated with diarrheal outbreaks in Guangdong, China (Ke et al., 2011). We found that both the serovars harbored the tdh virulence gene but not the trh gene. Though V. parahaemolyticus is an environmental organism, presence of tdh gene makes them virulent and hence detection of this marker gene is important (Meador et al., 2007). In our previous study, we found that both O3:K6 and O4:K8 serovars were identified as pandemic strains by various molecular typing techniques (Chowdhury et al., 2000). Interestingly, in this study, the two O4:K8 strains were negative in the GS-PCR, indicating the recent changes in the pandemic strains of V. parahaemolyticus at the molecular level.
Based on the surveillance of enteric pathogens conducted at the IDH, the serovars O3:K6 and O4:K8 harboring tdh identified in the present outbreak are not new in this region. Interestingly, the PFGE profile of an O3:K6 strain isolated from a diarrheal patient described in this report matched with a similar phenotypic strain isolated from a diarrheal case admitted during 2011 at the IDH. The occurrence of this O3:K6 strain indicates that there is a clonal pool of such strains persisting and circulating within the region. However, the pandemic V. parahaemolyticus O3:K6 serovar that was involved in the 2003 outbreak was different from the 2011 strains described in this report. On the other hand, O4:K8 strains isolated in the current outbreak and the IDH strain isolated in 2011 were clonally different.
V. parahaemolyticus strains are generally susceptible to commonly used antimicrobials in the treatment of diarrhea. We observed that except for ampicillin, all the tested strains were susceptible to all the antimicrobials. The patients administrated with the fluoroquinolone responded to the treatment, confirming the laboratory susceptibility results. Since the food samples were not microbiologically screened, we are not sure about the source of the V. parahaemolyticus encountered among diarrheal patients. Involvement of food-handlers cannot be ruled out since one of the early investigations conducted in this region has shown that the carrier status of V. parahaemolyticus is 21.4% (Sircar et al., 1979).
In summary, the finding described in this report shows the association of V. parahaemolyticus O3:K6 and O4:K8 serovars with an acute diarrheal outbreak. We suspect that this outbreak may have a link with consumption of contaminated food with V. parahaemolyticus. However, leftover foods were not available for in-depth investigation. Considering the nature of reoccurrence of foodborne outbreaks caused by V. parahaemolyticus in this region, the public health authorities ought to give due importance to these pandemic strains.
Footnotes
Acknowledgment
This investigation was supported in part by the Indian Council of Medical Research, and Indian Integrated Disease Surveillance Program, Ministry of Health and Family Welfare, New Delhi.
Disclosure Statement
No competing financial interests exist.