
Abstract
Cronobacter spp. (formerly Enterobacter sakazakii) is an emerging foodborne pathogen consisting of seven species including C. sakazakii, C. malonaticus, C. muytjensii, C. turicensis, C. dublinensis (with three subspecies, dublinensis, lausannensis, and lactaridi), C. universalis, and C. condimenti. To date, 12 Cronobacter serogroups have been identified. In this study, MboII restriction fragment length polymorphism patterns and DNA sequences of O-antigen gene clusters were used to identify novel serogroups of Cronobacter spp. Sequence analysis of the O-antigen regions, located between galF and gnd, of strains with distinct restriction fragment length polymorphism patterns revealed five unique gene clusters. These new O-antigen gene clusters were species specific and were termed C. turicensis O3, C. muytjensii O2, C. dublinensis O1, C. dublinensis O2, and C. universalis O1. Polymerase chain reaction assays were developed using primers specific to O-antigen processing genes and used to screen a collection of Cronobacter strains to determine the frequency of these newly identified serotypes.
Introduction
C
The O-antigen is a highly variable component of the lipopolysaccharide of Gram-negative bacteria and is used for the development of both serological and molecular typing methods. This structure of oligosaccharide repeats ranges in size from two to eight sugar residues, and its variability comes from the presence of different sugars, their unique order and arrangement, and the chemical bonds between residues (Samuel and Reeves, 2003). O-antigen encoding gene clusters are flanked by the galF and gnd genes in most Enterobacteriaceae, contain from six to 19 genes, and are usually 6- to 20-kb in length (Samuel and Reeves, 2003). Typically O-antigen gene clusters contain three types of genes: those specific to the biosynthesis pathways of nucleotide sugars, glycosyltransferase genes that provide unique linkages between sugar residues, and O-antigen processing genes required for assembly of polysaccharide repeat units and their subsequent transport through the periplasm (Samuel and Reeves, 2003). There are three known pathways for O-antigen processing, including one, which is Wzx/Wzy-dependent, an ABC transporter-dependent pathway, and one, which is synthase-dependent (Raetz and Whitfield, 2002; Cuthbertson et al., 2010). The most common O-antigen processing pathway in Enterobacteriaceae is the Wzx/Wzy-dependent transport process, which utilizes two proteins, an O-antigen flippase (Wzx), and an O-antigen polymerase (Wzy). However, in some instances O-antigens are processed via an ABC transporter mechanism using the Wzm protein in conjunction with an ATP binding protein, designated Wzt (Lewis et al., 2012; Cuthbertson et al., 2010).
The use of serotype-specific polymerase chain reaction (PCR) assays in lieu of the more traditional serological methods is accepted for typing Escherichia coli, Salmonella species, Shigella species, and many other Gram-negative bacteria including Cronobacter spp. (Wang et al., 2007; Liu et al., 2008; Mullane et al., 2008; Liu et al., 2010; Jarvis et al., 2011; Debroy et al., 2011). Previous work in our laboratory and others has identified 12 Cronobacter serogroups including C. sakazakii serogroups O1 through O7, C. malonaticus serogroups O1 and O2, C. muytjensii serogroup O1, and C. turicensis serogroup O1 and O2 (Mullane et al., 2008; Sun et al., 2011; Jarvis et al., 2011; Sun et al., 2012b; Sun et al., 2012a). In this study, we extend the characterization of Cronobacter O-antigen loci by adding five new serogroups. Furthermore, we developed five new serogroup-specific PCR assays to facilitate the characterization of Cronobacter spp.
Materials and Methods
Bacterial strains used in this study
A collection of 270 bacterial isolates, representing seven Cronobacter species, was used in this study. All strain information and results were submitted to the Pathogen Annotated Tracking Resource Network, located at
Restriction fragment length polymorphism (RFLP) analysis of O-antigen amplicons by use of MboII
O-antigen gene clusters ranging in size from 9794 to 14,788 bp were amplified using primers specific to the galF and gnd genes of Cronobacter (Table 2) as described previously (Jarvis et al., 2011). Amplicons recovered (1.5 μg) were digested with MboII (New England BioLabs, Ipswich, MA) according to the manufacturer's instructions. Restriction digests were subjected to gel electrophoresis using 1.5% agarose gels and were visualized with ethidium bromide (0.5 μg/mL, final concentration). TIFF images were imported into BioNumerics software (version 6.6, Applied Maths, Inc., Austin, TX), and dendrograms were generated using the Dice similarity coefficient and the unweighted-pair group method with arithmetic means method. A band tolerance of 1% and optimization coefficient setting of 3% were applied.
This primer pair was used to amplify the O-antigen regions of Cronobacter (C.) muytjensii O2, C. turicensis O3, and C. universalis O1.
Amplification and sequencing of Cronobacter O-antigen gene clusters
The O-antigen regions of five Cronobacter strains were mined from annotated whole genome sequences in RAST (Aziz et al., 2008) using the SEED viewer (Overbeek et al., 2005). The Whole Genome Shotgun project has been deposited at DDBJ/EMBL/GenBank and the strain and accession numbers for the genome sequences are Cronobacter dublinensis subsp. lactaridi LMG 23825T (AJKX00000000), Cronobacter dublinensis subsp. lausannensis LMG 23824T (AJKY00000000), Cronobacter dublinensis subsp. dublinensis LMG 23823T (AJKZ00000000), Cronobacter muytjensii ATCC 51329T (AJKU0000000), Cronobacter universalis NCTC 9529T (AJKW00000000). When it was necessary, fragmented gene clusters were assembled using BioNumerics software version 6.6 (Applied Maths, Inc., Austin, TX). BLASTp analysis was used to predict open reading frames (ORFs) and in cases where this analysis indicated translation errors or when gaps were detected, further sequencing was performed as described previously (Jarvis et al., 2011). The O-antigen gene cluster of Cronobacter turicensis E609 was amplified using primers specific to the galF and gnd genes of Cronobacter as described above and sequenced using the primer walking method (Giesecke et al., 1992). The bacterial genomic DNA purification and PCR amplification methods were as described above and the DNA was submitted to Eurofins MWG Operon (Ebersberg, Germany) for sequencing.
Analysis of Cronobacter O-antigen gene clusters
The Pfam database was used to search for conserved protein domains, and phylogenetic analyses were done within MEGA5 (Finn et al., 2008, Tamura et al., 2011). Evolutionary relationships of wzx and wzy nucleotide sequences were determined within MEGA5 (Tamura et al., 2011), using the neighbor-joining method (Saitou and Nei, 1987), with evolutionary distances computed using the maximum composite likelihood method (Tamura et al., 2004). Prediction of transmembrane-spanning domains of Wzx and Wzy were identified using the Hidden Markov Models (TMHMM) program (Krogh et al., 2001).
Development of serogroup-specific PCR assays
PCR primers specific to the wzx genes of C. turicensis E609, C. dublinensis subspecies lactaridi LMG 23825T, C. dublinensis subspecies dublinensis LMG 23823T, C. muytjensii ATCC 51329T, C. universalis NCTC 9529T, and the wzm gene of C. dublinensis subspecies lausannensis LMG 23824T were designed (Table 2). Table 2 also contains primer sequences for the wzx gene from C. sakazakii O4 strain E764 that was part of this study. Simultaneous to this work, another group characterized this O-antigen gene cluster and developed a serogroup assay specific to the wzy gene of C. sakazakii O4 (Sun et al., 2012b). The PCR cycling conditions for all reactions used to screen 270 strains included an initial denaturation step at 95°C for 2 min, followed by 25 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 1 min, with a final extension step at 72°C for 5 min.
Nucleotide sequence accession numbers
The nucleotide sequences of the O-antigen regions for C. turicensis E609, C. dublinensis subspecies lactaridi LMG 23825T, C. dublinensis subspecies dublinensis LMG 23823T, C. dublinensis subspecies lausannensis LMG 23824T, C. muytjensii ATCC 51329T, C. universalis NCTC 9529T, and C. sakazakii O4 E764 were deposited in GenBank under accession numbers JX475926, JQ390549, JQ390550, JQ390551, JQ390552, JQ390553, and JQ390548, respectively.
Results
RFLP cluster analysis of Cronobacter O-antigen gene clusters with MboII restriction digestion
RFLP analysis was performed on a subset of Cronobacter spp., including the strains from which the O-antigen regions were sequenced, revealing five novel Cronobacter molecular-based serogroup clusters (Fig. 1). Two RFLP clusters were identified for C. dublinensis, one for C. muytjensii, a single cluster for C. turicensis, and the RFLP pattern of the single C. universalis strain is distinct from those of all of the other strains (Fig. 1). One of the C. dublinensis clades contains three distinct RFLP patterns and the C. turicensis clade consists of two patterns (Fig. 1).
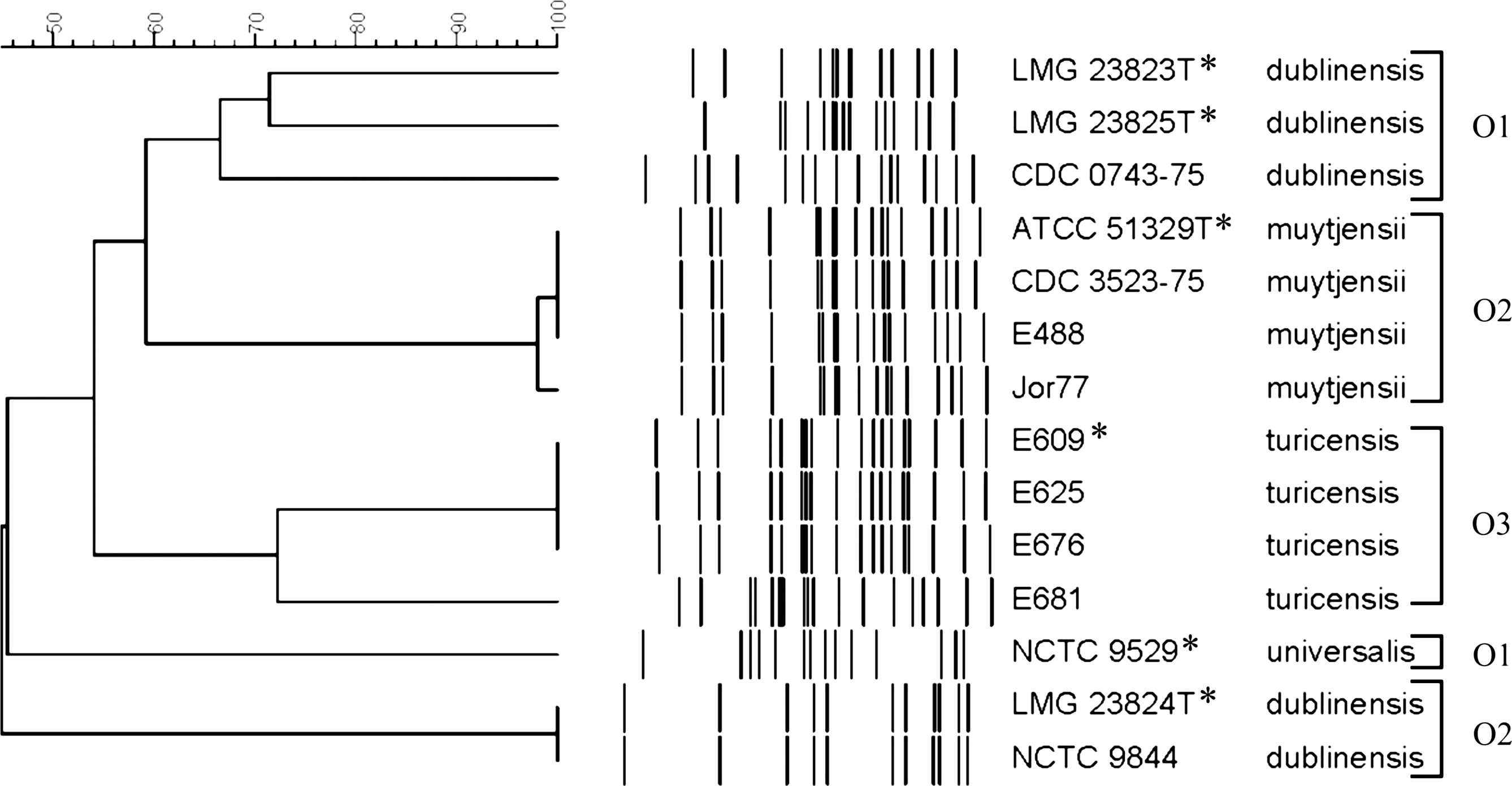
Restriction fragment length polymorphism analysis of MboII digests from Cronobacter O-antigen gene clusters. The dendrogram was generated using Bionumerics software with an 80% similarity value used to distinguish serogroup specific clusters. An asterisk (*) indicates the Cronobacter strains analyzed in this study. The scale at the top represents percent similarity.
Sequence analysis of novel Cronobacter serogroup-specific O-antigen gene clusters
The nucleotide sequences from the O-antigen gene clusters (including the galF and gnd genes) of C. turicensis E609, C. dublinensis subspecies lactaridi LMG 23825T, C. dublinensis subspecies dublinensis LMG 23823T, C. dublinensis subspecies lausannensis LMG 23824T, C. muytjensii ATCC 51329T, and C. universalis NCTC 9529T were 13, 275, 12,883, 13,867, 9,794, 14,788, and 12,288 bp in length, respectively. BLASTp analysis revealed five unique O-antigen gene clusters among these six strains (Table 3 and Fig. 2).
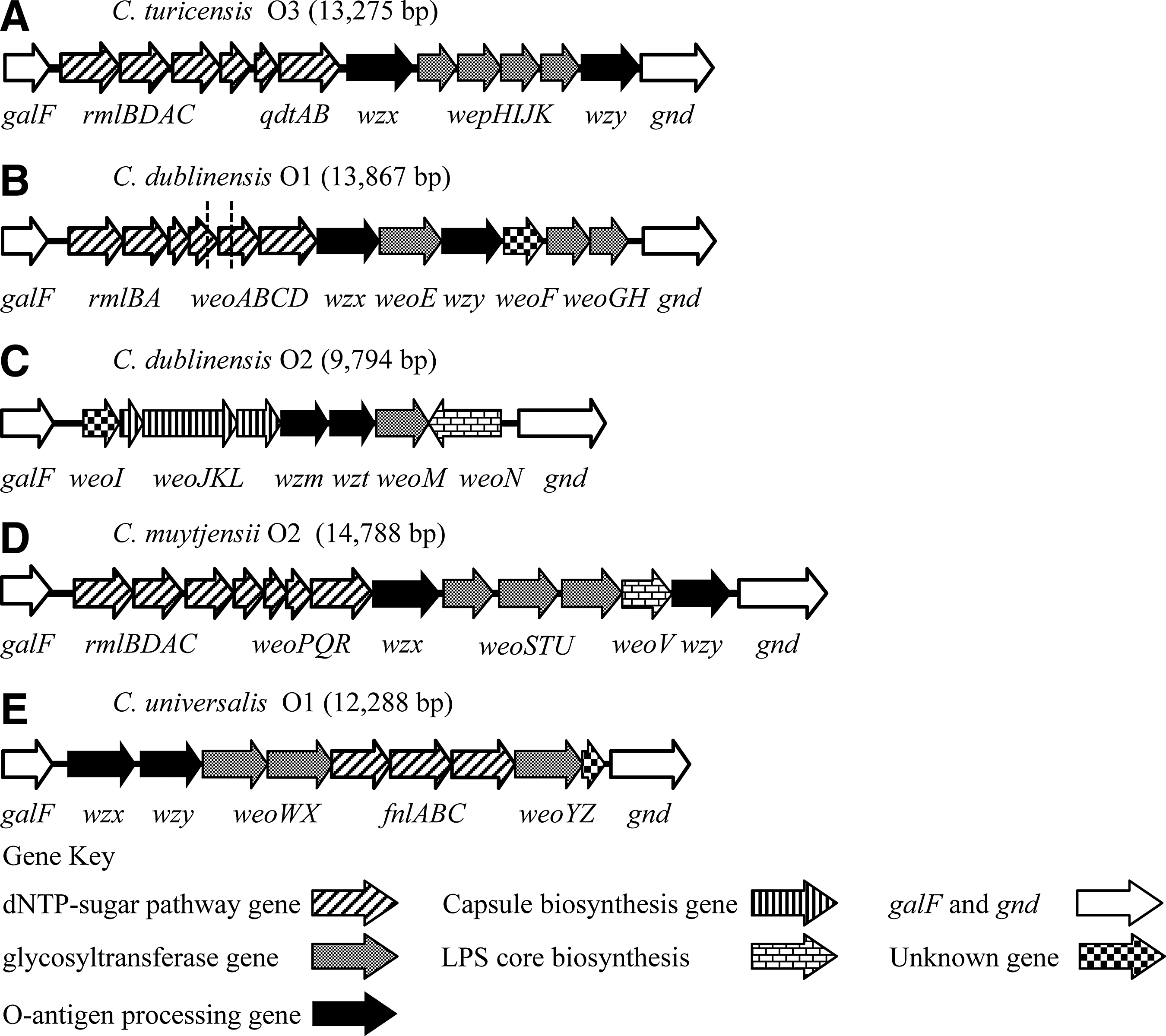
O-antigen gene clusters of Cronobacter serogroups.
ORFs, open reading frames.
The O-antigen gene cluster of C. turicensis E609 was designated as serogroup C. turicensis O3 and consists of 12 predicted proteins, 11 of which are 81–99% homologous to C. sakazakii serogroup O5 (Table 3 and Fig. 2). The nucleotide sequence of C. dublinensis LMG 23825T and LMG 23823T O-antigen gene clusters, designated as serogroup C. dublinensis O1, are 95% similar with one another and they encode for the same 12 ORFs (Table 3 and Fig. 2). A second C. dublinensis serogroup, C. dublinensis O2, was identified in strain LMG 23824T. This gene cluster encodes for eight predicted proteins that share 42–82% homology with a predicted polysaccharide synthesis region in Pantoea ananatis and both of these O-antigen regions encode for predicted ABC transporter genes for O-antigen processing (Table 3 and Fig. 2).
The O-antigen gene cluster from C. muytjensii ATCC 51329T, designated serogroup C. muytjensii O2, encodes for 13 predicted proteins. ORFs 1–5 were 75–100% homologous to the predicted proteins of the C. muytjensii serogroup O1 gene cluster (Jarvis et al., 2011). ORFs 7–13 in this gene cluster were 58–89% homologous to O-antigen proteins from C. sakazakii O5 (Table 3 and Fig. 2) (Jarvis et al., 2011; Sun et al., 2012b).
Finally, the O-antigen gene cluster of C. universalis NCTC 9529T, designated C. universalis serogroup O1, encodes nine predicted proteins. ORF 1 and ORFs 3–6 encode for predicted proteins that were 78–98% homologous to proteins in the C. sakazakii O7 O-antigen region (Sun et al., 2012b). ORF 2 of C. universalis O1, a feature that was lacking from the C. sakazakii O7, encodes for an O-antigen polymerase, Wzy, and was 43% homologous to the Wzy protein from E. coli O98:H8 (Table 3). The last three predicted proteins in the C. universalis O1 gene cluster share 95–97% homologies with the last three ORFs predicted in the O-antigen region of C. turicensis serogroup O1 (Table 3) (Jarvis et al., 2011).
Nucleotide sugar synthesis pathway genes
BLASTp analysis of the C. turicensis O3 O-antigen gene cluster revealed six predicted sugar synthesis proteins including RmlBDAC, and QdtA and QdtB. QdtC, which is usually present in conjunction with QdtA and B, is missing in the C. turicensis O3 O-antigen region because of a 396-bp gap in the nucleotide sequence revealed by BLASTn analysis of C. turicensis O3 and C. sakazakii O5 (Table 3 and Fig. 2) (Sun et al., 2012b). The C. dublinensis O1 gene clusters from LMG 23825T and LMG 23823T contain six predicted sugar synthesis genes. However, there was a 984-bp deletion in LMG 23823T, resulting in truncations in the ORFs encoding WeoB, which is predicted to encode for an aceytltransferase, and WeoC, which has no predicted function (Table 3 and Fig. 2). The C. dublinensis O2 O-antigen gene cluster does not contain any predicted sugar synthesis genes, suggesting that the sugars in this oligosaccharide structure may be coded for elsewhere in the genome (Table 3 and Fig. 2). The O-antigen gene cluster from ATCC 51329T, designated as C. muytjensii O2, encodes for seven predicted nucleotide sugar synthesis proteins, including RmlBDAC enzymes involved in the synthesis of dTDP-L-rhamnose (Samuel and Reeves, 2003). These four ORFs, as well as ORFs 5, 6, and 7 of this gene cluster, share identity with ORFs 1–7 of the C. muytjensii O1 gene cluster, suggesting a similar sugar repeat oligosaccharide structure for these two serogroups (Table 3 and Fig. 2) (Jarvis et al., 2011). The O-antigen gene cluster of C. universalis NCTC 9529T serogroup O1 encodes for three putative sugar synthesis proteins, including FnlABC, which catalyzes the synthesis of 2-acetamido-2,6-dideoxy-hexose (UDP-L-FucNAc) in E. coli and Shigella O-antigens (Table 3 and Fig. 2) (Samuel and Reeves, 2003; Perepelov et al., 2006; Liu et al., 2008).
Glycosyltransferase genes
Twenty-six families of glycosyltransferase genes have been identified based on amino acid homology (Campbell et al., 1997). The O-antigen gene cluster of C. turicensis O3, C. dublinensis O1, C. dublinensis O2 gene clusters encode for four, three, and one predicted glycosyltransferase proteins, respectively (Table 3 and Fig. 2). The C. muytjensii serogroup O2 O-antigen region possesses the genes for two glycosyltransferases: one each from family 1 and family 2 (Table 3 and Fig. 2). Finally, there are three glycosyltransferases predicted from the O-antigen gene cluster of C. universalis serogroup O1 (Table 3 and Fig. 2).
O-antigen processing genes
The O-antigen gene clusters of C. turicensis O3, C. dublinensis O1, C. muytjensii O2, and C. universalis O1 all encode predicted Wzx and Wzy proteins (Table 3 and Fig. 2). TMHMM analysis showed that all of the predicted Wzx proteins have between eight and 13 transmembrane-spanning domains and the Wzy proteins were predicted to have between eight and 10 transmembrane-spanning domains. BLASTp analysis of the predicted O-antigen processing proteins from the C. dublinensis O2 gene cluster revealed that they utilize ABC-transporter proteins for O-antigen processing. In comparison to the O-antigen gene cluster of P. ananatis strain AJ13355, ORFs 5 and 6 are 81% and 61% homologous to Wzm and Wzt, respectively (Table 3 and Fig. 2) (Hara et al., 2012). The Wzm protein is an ABC-2 type transporter permease, and the Wzt protein is an ABC O-antigen transporter (Table 3 and Fig. 2).
Serogroup-specific PCR assays for Cronobacter
Analysis of the Cronobacter O-antigen regions sequenced in this study facilitated the design of six serogroup-specific assays. Five of the assays use PCR primers specific to the O-antigen wzx gene sequences (namely, C. turicensis O3, C. sakazakii O4, C. dublinensis O1, C. muytjensii O2, and C. universalis O1) (Table 2). A sixth serogroup-specific assay was designed using primers specific to the wzm gene identified in the C. dublinensis O2 gene cluster (Table 2). Table 4 summarizes previous results (Jarvis et al., 2011; Mullane et al., 2008) along with those obtained using these new PCR assays to screen our strain collection of 270 Cronobacter isolates. The C. turicensis O3 PCR assay identified a new group of five (2%) strains in the collection. Sixteen (6%) C. sakazakii O4 strains and all six (2%) C. dublinensis O1 strains in the collection were positively identified by wzx PCR assays, and the C. dublinensis wzm assay was positive with four serogroup O2 strains. The new C. muytjensii serogroup O2 consists of five (2%) strains, and the C. universalis O1 serogroup is represented by one strain in our collection (Table 4).
PCR specific to the wehC gene (Mullane et al., 2008).
PCR specific to the wehI gene (Mullane et al., 2008).
PCR specific to the wzx gene (this study and (Jarvis et al., 2011)).
PCR specific to the wzm gene (this study).
ND, not determined (negative with Cronobacter PCR assays developed in this and previous studies).
These data combined with the RFLP data in Figure 1 revealed several novel serogroup-specific Cronobacter clades; all of them were species-specific (Table 4 and Fig. 1). It was of interest that C. sakazakii strains in the present study and previously reported as an undetermined serogroup (Jarvis et al., 2011) were identified as serogroup O4 using a wzx PCR assay (Table 4). These results correlate well with those reported by Sun et al. (Sun et al., 2012b), who described a C. sakazakii serogroup O4 strains identified using a wzy-based PCR assay, which is also positive for the same group of strains. Twenty-five (9%) of the isolates in our collection were negative with the Cronobacter serogroup-specific PCR assays developed in this and previous studies (Table 4) (Mullane et al., 2008; Jarvis et al., 2011). Of these, there are 15 C. sakazakii, two C. turicensis, five C. muytjensii, one C. dublinensis, one C. universalis, and one C. condimenti PCR-negative strains suggesting additional serogroups in the genus.
Discussion
In this study, our BLASTp analysis of Cronobacter O-antigen gene sequences revealed several predicted and shared proteins between bacterial genera and species. Predicted proteins of the C. muytjensii O2 O-antigen region share homologies with the C. muytjensii O1 and C. sakazakii O5 O-antigen regions (Table 3) (Jarvis et al., 2011). The results from predicted proteins (Table 3) and nucleotide phylogeny (Fig. 3) suggest that C. turicensis O3 and C. sakazakii O5 have descended from a common ancestor that was also closely related to C. muytjensii O3, a prediction that is supported by multilocus sequence type (MLST) analysis (Joseph et al., 2012). The wzx and wzy nucleotide phylogeny of the C. sakazakii O5, C. muytjensii O2, and C. turicensis O3 clade with its closest neighbor, the C. sakazakii O3 and C. muytjensii O1 clade, supports the overlap in predicted proteins from the O-antigen genes clusters of these two groups (Fig. 3 and Table 3) (Jarvis et al., 2011). The first six predicted proteins in the C. universalis O1 region are homologous with C. sakazakii O7 proteins, with the exception of ORF 2, which is missing, in C. sakazakii O7 (Table 3) (Sun et al., 2012b). Phylogenetic analysis of the nucleotide sequences of wzx and wzy genes confirms this close relationship between these serogroups (Fig. 3 and Table 3). The nesting of the E. coli O103 strain with the C. sakazakii O4 strains reflects a shared antigenic structure between the two species at the genetic level (Fig. 3). The C. dublinensis O2 gene cluster and the O-antigen gene cluster from P. ananatis demonstrated 45–82% protein homology and seven of the eight predicted proteins in this gene cluster were functionally similar; the exception is ORF 2, which is homologous to a predicted capsule biosynthesis protein from Zymomonas mobilis subsp. mobilis (Table 3). Additionally, they both encode for ABC-transporter genes for O-antigen processing. These data suggest that the O-antigen gene clusters of P. ananatis and C. dublinensis O2 have a similar evolutionary origin. The proteins with the highest homology in these two O-antigen regions are Wzm and Wzt, the O-processing genes, which are usually the most divergent genes in O-antigen regions containing ABC-transporter genes (Cuthbertson et al., 2010).
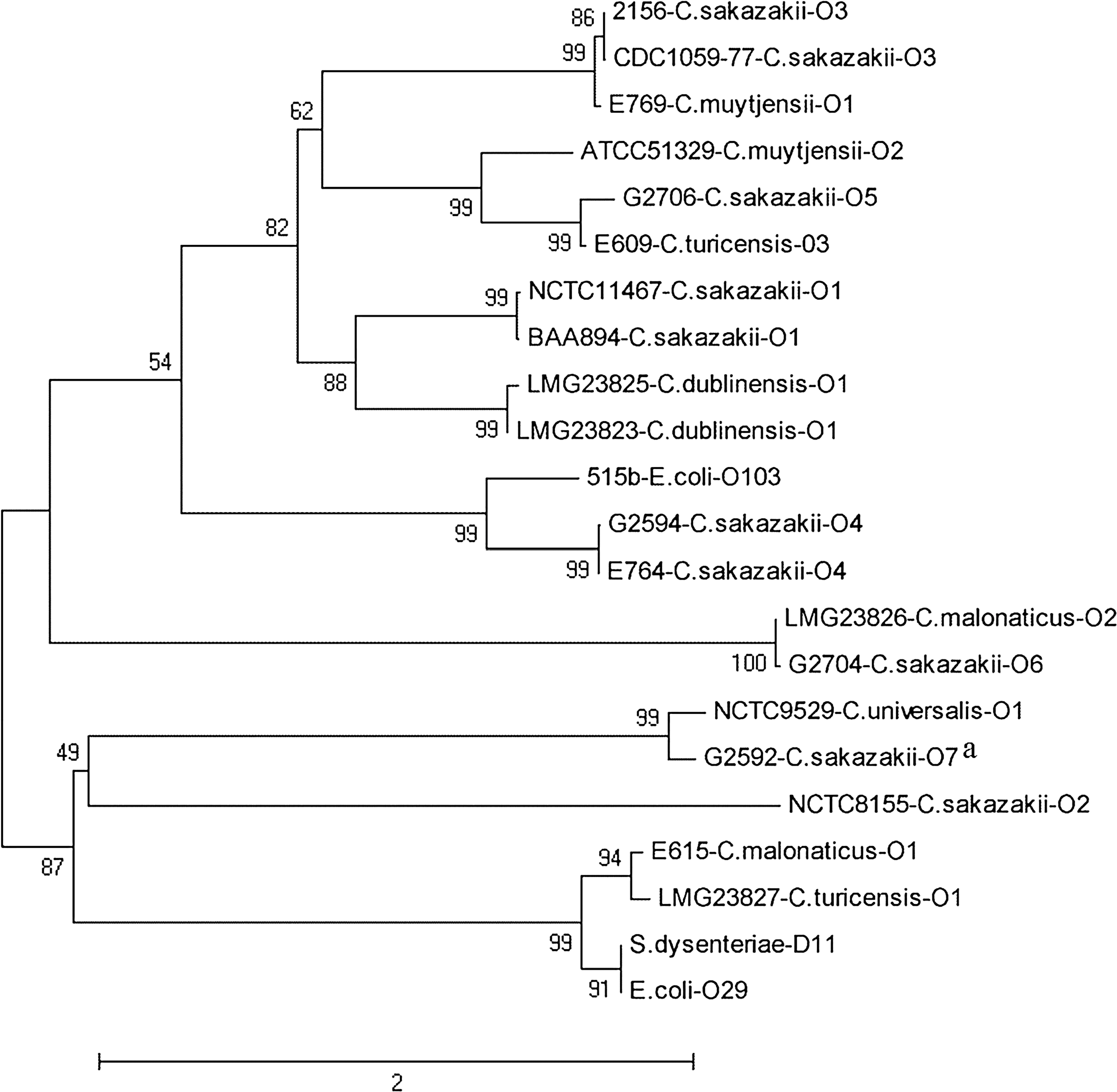
Evolutionary relationships of concatenated wzx and wzy nucleotide sequences from Cronobacter serogroups, Escherichia coli O29, E. coli O103, and Shigella dysenteriae D11. Relationships were analyzed in MEGA5 using the Neighbor-Joining method with distances computed using the Maximum Composite Likelihood method. Numbers on the branches denote bootstrap percentages from 1000 bootstrap replicates with a cut-off value of 50%. The accession numbers for the O-antigen gene clusters are as follows: Cronobacter NCTC 11467 (ATCC 29544T) (EU076545), BAA 894 (NC_009778), NCTC 8155 (EU076546), G2594 (JQ674747), G2706 (JQ674748), G2704 (JQ674749), G2592 (JQ674750), E. coli O103 (EF027106), S. dysenteriae D11 (EU294172), and E. coli O29 (EU294173). aThe nucleotide sequence from C. sakazakii O7 G2592 does not contain wzy.
Not surprisingly, we identified two serogroups in this study, C. dublinensis O1, and C. turicensis O3, that consist of strains that were PCR-positive despite their vastly different RFLP fingerprints (Fig. 1 and Table 3). This result, also observed in previous work by Jarvis et al. (Jarvis et al., 2011), indicates that although RFLP analysis of O-antigen gene clusters has proven to be useful for initial screening, different patterns do not always correlate with different O-antigens. Further serogroup-specific PCR amplification is needed for the confirmation in this case.
Conclusions
In conclusion, we identified five new Cronobacter spp. serogroups using PCR assays specific to O-antigen genes. Four of these PCR assays are specific to wzx genes in C. turicensis O3, C. muytjensii O2, C. dublinensis O2, and C. universalis O1. One assay is specific to the wzm gene found in the O-antigen gene cluster of C. dublinensis O2 strains, which have ABC transporter mechanisms for O-antigen processing. Unlike the PCR assays previously described for Cronobacter, these new assays are all species-specific and represent the continuation of an approach that is useful for monitoring Cronobacter serotypes found among clinical, environmental, and food isolates.
Footnotes
Acknowledgments
We thank Atin R. Datta, Barbara A. McCardell, and Kevin Gaido, for reviewing this manuscript. K. G. Jarvis, L. Hu, and C. J. Grim are Oak Ridge Institute for Science and Education fellows and the authors wish to thank the Department of Energy for their support.
Disclosure Statement
No competing financial interests exist.