
Abstract
Antibiotic growth promoters (AGPs) have been used as feed additives to improve average daily weight gain and feed efficiency in food animals for more than five decades. However, use of AGPs is associated with the emergence of antibiotic-resistant human pathogens of animal origin, posing a significant threat to food safety and public health. Thus, development of novel alternatives to AGPs is important to mitigate antimicrobial resistance in foodborne pathogens. To achieve this goal, the mode of action of AGPs should be elucidated. In this study, the response of the chicken gut microbiota to AGPs was examined using two culture-independent approaches: phospholipid fatty acid (PLFA) biomarker analysis and 16S rDNA clone library sequencing. PLFA analysis showed that AGP tylosin treatment changed composition of the microbiota in various intestinal sites; however, total viable bacterial biomass in intestine was not affected by tylosin treatment. PLFA analysis also revealed an abundant viable fungal population in chicken microbiota. Eight 16S rDNA libraries (96 clones per library) were constructed using ileal samples from chickens that received either antibiotic-free or medicated feed. The 16S rDNA clone analysis of the growth-relevant samples showed the AGP treatment influenced the diversity of ileum microbiota in the chickens primarily in the Firmicutes division. In particular, Lactobacillus spp. populations in the ileum of AGP-treated chickens were significantly lower than those from chickens receiving antibiotic-free feed. Together, this study revealed novel features of the intestinal microbiota in response to AGP treatment and suggested approach to develop potential alternatives to AGPs for mitigation of antimicrobial resistance in foodborne pathogens.
Introduction
A
Understanding the mode of action of AGPs is critical for developing effective and novel AGP alternatives. Given the antibacterial nature of AGPs and important role of intestinal microbiota in host nutrient assimilation and immune defense, it is widely accepted that the growth promoting effect of AGPs is primarily mediated through enhanced energy harvest and the control of gastro intestinal infections due to altered microbiota in the intestine (Backhed et al., 2005; Dibner and Richards, 2005). However, the precise mechanisms of AGPs are still largely unknown. It has been proposed that AGPs may improve animal growth performance by reducing the maintenance cost of gastrointestinal tract system via reduction of total gut bacteria (Collier et al., 2003; Gaskins et al., 2002) and/or by reshaping the microbial diversity and relative abundance in the intestine to achieve optimal microbiota for growth (Dibner and Richards, 2005). Several products, such as organic acids, probiotics, and prebiotics, have been used to alter the intestinal microbiota for improving animal health and production (Dibner and Richards, 2005). However, very limited data is available to justify the choice of specific bacterial species or products for such microbiota manipulation. Thus, examination of microbial diversity and dynamics in response to AGP treatment could provide key information for the development of novel alternatives to AGPs.
With the aid of 16S rDNA-based culture-independent approaches, investigations on the effect of AGPs on intestinal microbiota have been studied in different food animals, including poultry and swine (Collier et al., 2003; Danzeisen et al., 2011; Dumonceaux et al., 2006; Kim et al., 2012; Knarreborg et al., 2002; La-pngkhum et al., 2011; Wise and Siragusa, 2007), which have greatly improved our understanding of intestinal microbiota changes in response to AGPs. However, these studies did not address if AGP usage affected total viable bacteria in the intestine as proposed previously (Collier et al., 2003; Gaskins et al., 2002). In addition, these studies either failed to show the growth-promoting effect of AGPs or lacked any growth/nutritional measurements in conjunction with microbial ecology analysis, which reduced the relevance of the observed microbial structure shift to animal growth. Therefore, to examine the complex response of gut microbiota to AGP treatment, innovative approaches as well as high-quality, growth-relevant intestinal samples should be used.
In this study, a unique phospholipid fatty acid (PLFA) biomarker analysis was used to determine viable biomass and composition of gut microbiota in chickens subjected to long-term treatment with AGP tylosin. A chicken experiment was further performed with typical AGPs (salinomycin, bacitracin, and virginiamycin) under similar environmental conditions in poultry industry. The growth-relevant ileal samples were further used for construction of 16S rDNA clone libraries and corresponding microbial ecology analysis.
Materials and Methods
Animal experiments and intestinal sample collections
Experiment 1
The samples used for PLFA analysis were derived from a previous chicken experiment which determined if long-term use of tylosin (a macrolide AGP) selects for macrolide-resistant Campylobacter in chickens (Lin et al., 2007). Upon termination of the chicken experiment at 41 days of age, two chickens were randomly selected from control and tylosin-treated group, respectively, for sample collection. The mid-jejunum, mid-ileum, and cecum sections were removed from each chicken and were immediately stored on dry ice on site and then stored at −80°C prior to PLFA analysis.
Experiment 2
To obtain high-quality intestinal samples suitable for studying diversity and abundance of gut microbiota in response to AGP treatment, a well simulated chicken experimental system was established using conditions similar to those used for raising poultry in the industry. Briefly, 300 one-day-old Hubbard broiler chicks were randomly allotted to 20 floor pens of 15 chicks each. Each floor pen had at least 12.5 square feet floor space. The room was maintained at a minimum of 24°C, and gas brooders were used to provide supplemental heat through day 14. A 23-h light/1-h dark photoperiod was used through week 2, followed by an 18-h light/6-h dark photoperiod for the remainder of the experiment. To simulate growth conditions on the litter and ensure chicks were exposed to diverse normal intestinal flora for observation of beneficial growth response to AGPs, pens contained new softwood shavings top-dressed with about 1/4” to 1/2” of used litter from a commercial broiler farm, which is a standard practice in the United States for poultry industry to achieve better performance. This practice also has been used successfully to replicate industrial observations in experimental systems (Waldroup et al., 2005).
Corn and soybean meal based diets were formulated to meet current industry standards. A single batch of each diet type (starter, grower, and finisher) were mixed and then divided in half to prepare the two experimental diets (medicated and non-medicated), thus eliminating any variability in diet nutrient composition. The medicated starter and grower diets were supplemented with salinomycin and bacitracin methylene disalicylate (each at 0.50 g/kg feed) and the medicated finisher diets were supplemented with virginiamycin (at 10 g/ton feed). Non-medicated diets were served as the control. The non-medicated and medicated diet series were randomly assigned to 10 pens each. Water and feed were provided ad libitum. Body weight was determined on days 7, 14, 21, 32, and 42. Effect of diet treatment (medicated vs. non-medicated) on body weight gain was assessed using SAS mixed model analysis of variance (SAS Institute, Cary, NC). Differences were considered significant when p≤0.05. At days 14, 32, and 42, one bird from each pen whose body weight was nearest the mean for the pen was removed for intestinal sample collection. The ileal contents were immediately transferred to sterile cryogenic vials, frozen on dry ice, and then stored at −80°C prior to analysis.
PLFA analysis
PLFA analysis offers exceptional ability to distinguish live from dead organisms and has been widely used to study live biomass and composition of complex microbial consortia in different environments (Pinkert et al., 2002). In this study, individual intestinal samples from two chickens in each treatment group were used for PLFA analysis. Whole intestinal samples (mucosa plus luminal contents) were lyophilized, weighted, and extracted using the modified one-phase chloroform-methanol-buffer system (White and Ringelberg, 1998). The total lipid extract was then fractionated into neutral lipids, glycolipids, and polar lipids by silicic acid column chromatography (Guckert et al., 1985). The polar lipid fraction was transmethylated to form the fatty acid methyl esters in mild alkali as described previously (Guckert et al., 1985). The fatty acid methyl esters were analyzed and identified by gas chromatography/mass spectroscopy using an Agilent 6890 series gas chromatograph interfaced to an Agilent 5973 mass selective detector with a 50-m non-polar column (0.2 mm I.D., 0.11 μm film thickness) with a temperature program of 100°C initial temperature, 10°C min−1 to 150°C for 1 min, 10°C min−1 to 282°C for 5 min with injector temperature at 270°C and detector at 290°C. Total analysis time was 55 min. All solvents used were of GC grade and were obtained from Fisher Scientific.
16S rDNA clone library construction and analysis
Total of eight libraries (four for medicated group and four for control group; 96 clones/library for each chicken) were constructed for the study. Briefly, total DNA was extracted directly from ileal content using ExtractMaster™ Fecal DNA Extraction Kit (Epicentre Biotechnologies, Madison, WI). The 16S rDNA cloning and sequencing were performed as described previously (Dumonceaux et al., 2006; Eckburg et al., 2005; Layton et al., 2000) with slight modifications. Briefly, the 16S rDNA were amplified from each chicken ileal DNA using broad-range eubacterial primers 530f (5’-GTGCCAGCMGCCGCGG-3’) and 1392r (5’-ACGGGCG GTGTGTRC-3) (Lane, 1991). The PCR products were purified and cloned into vector pGEM-T Easy (Promega, San Luis Obispo, CA). Ligation mixtures were used to transform Escherichia coli JM109 (Promega) according to manufacture's instruction. To construct library for each individual chicken, 96 white colonies were randomly picked into individual wells of a 96-well plate. Thus, total 768 plasmids were purified from transformants with Qiagen Miniprep Kit (Qiagen, Valencia, CA) and subjected to 16S rDNA sequencing using SP6 and T7 primers. The bidirectional sequence reads of the clone inserts provided 16S bacterial rDNA sequence of about 750 bp usable for phylogenetic analysis. The sequences were aligned to the small subunit rRNA ribosomal Database Project (RDP-II) (Cole et al., 2009). All chimeric sequences, eukaryotic (e.g., chicken) sequences, vector sequences, and sequence of poor quality were deleted from further phylogenetic analysis. The sequences were grouped into phylotypes using Olsen-corrected similarity matrices so that the least similar pair within the phylotypes shared at least 99% similarity, as suggested by Eckburg et al. (2005).
Results
Effect of tylosin treatment on bacterial biomass in the chicken intestine
The bacterial biomass was determined to include the total PLFA minus the normal saturates over 18 carbons in length and the polyenoic PLFA, which are generally associated with eukaryote biomass (Bossio and Scow, 1998; Harwood and Russell, 1984; White et al., 1997). The trace quantities of PLFA of unknown structure were also excluded from the prokaryote biomass. Based on the total bacterial biomass analysis (Fig. 1A), cecal and jejunal sections showed comparable bacterial biomass, which is slightly higher than ileal bacterial biomass in chickens treated with or without tylosin. However, there was no significant difference observed between treated and untreated samples (p>0.05).
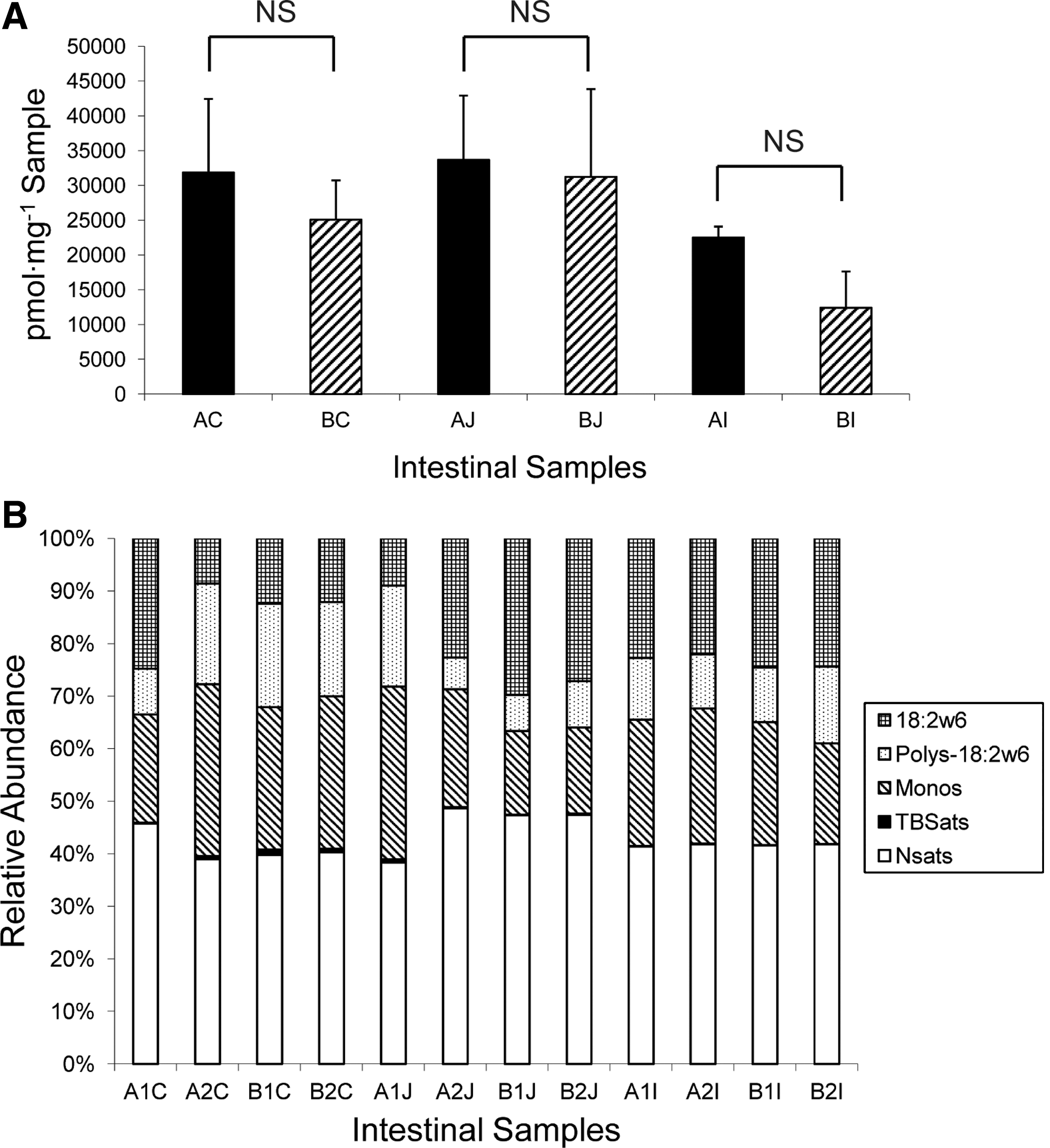
Phospholipid fatty acid (PLFA) analysis of gut microbiota of chickens receiving antibiotic-free feed (control group) or tylosin-containing feed (treatment group).
PLFA profile of intestinal microbiota
PLFA analysis revealed a relative simple community structure, primarily consisted of normal saturates, monounsaturates, polyunsaturates, and some terminally branched saturates (Fig. 1B). The eukaryote PLFA18:2ω6, which is a predominant PLFA in many fungi but is not found in native animal intestinal cells (Frostegard et al., 1993; Harwood and Russell, 1984; Pennanen et al., 1996), was detected among all samples with relative higher abundance in the ileum and jejunum. The total polyunsaturates, an indicator of eukaryote, showed an increase in jejunum samples from tylosin-treated chickens, which was largely due to the increase of PLFA 18:2ω6 in jejunum in response to tylosin (Fig. 1B). Notably, the long chain fatty acids over 18 carbons, which are associated with eukaryote biomass and were excluded for fungal biomass estimation in this study, may also be detected in some fungi (Harwood and Russell, 1984; Wells et al., 1996). Therefore, the relative abundance of total fungal biomass in the chicken intestine may be higher than the already unexpectedly high-level of fungi revealed by the PLFA analysis here.
With respect to bacterial populations, the community structures recovered from ileum section were stable and similar among treated and untreated samples. However, monounsaturates, the indicative of a large population of Gram-negative bacteria, showed a notable decrease in the treated jejunum section compared with the untreated jejunum section. This observation is consistent with the finding that tylosin treatment exerted a higher stress on Gram-negative bacteria in jejunum than in other sections as assessed from the ratios of the trans/cis PLFA, an indicator showing the physiological status of Gram-negative bacteria (data not shown). Terminally branched saturates that are often associated with Gram-positive bacteria and Gram-negative sulfate reducing bacteria showed a higher proportion in the cecum compared with that in the jejunum and ileum.
16S rDNA analysis of gut microbiota in response to AGP treatment
In response to AGP treatment, the medicated chickens grew significantly more slowly during the first week and eventually caught up with the control birds by week 2 and showed signs of increased growth during grower phase (days 14–32) (Table 1). By day 42, body weight gain in the medicated group was significantly higher (p<0.05) than that in control group, which is consistent with the observations in poultry industry.
p≤0.05.
The data consists of body weight means with standard deviations from 10 pens and are expressed on a bird basis. The levels of statistical significance between –AGP and+AGP were determined for each time point.
The 16S rDNA clone libraries were made individually for each ileal sample from four chickens in AGP-untreated (control) or AGP-treated (medicated) groups. A total of 768 clones were selected and plasmids from these clones were subjected to sequencing and phylogenetic analysis. As shown in Table 2, the AGP treatment influenced the diversity of ileum microbiota in the chickens primarily in the Firmicutes division. The four individual chickens in each treatment displayed similar microbiota pattern in terms of type and abundance of bacterial groups (data not shown). Notably, AGP treatment eliminated Lactobacillus spp in each of AGP-treated chickens while Lactobacillus spp still accounted for a large proportion of the microbiota of the ileum in the control chickens.
Discussion
This work confirmed previous findings that the composition of intestinal microbiota was influenced by AGP treatment (Collier et al., 2003; Dumonceaux et al., 2006; Knarreborg et al., 2002; Wise and Siragusa, 2007). However, several unique features make this pilot study distinct from previous microbiota work. First, this study utilized a unique and complementary molecular technique (i.e., PLFA) to examine complex intestinal ecosystems including both mucosal tissue and luminal contents, which led to some novel findings. Second, this study successfully simulated the conditions used for raising poultry in industry and observed the growth-promoting effect of AGPs in an experimental system. Establishment of this experimental system provides us high quality, growth-relevant intestinal samples for examining the effect of AGPs on gut microbiota. Finally, using the growth relevant ileal samples, the pilot 16S rDNA analysis in this study suggested that the lactobacilli population is actively involved in host metabolism and energy harvest.
The 16S rDNA-based molecular approaches have been widely and successfully used to study intestinal microbiota. However, one of limitations of the 16S rDNA-based approaches is their inability to distinguish live from dead organisms. The live biomass of complex microbial consortia can be determined by measuring the concentrations of the universal components of cell membranes, such as the lipids that serve as biomarker “signatures” for specific groups of microbes (White and Ringelberg, 1998). Phospholipids have an active metabolism and do not persist after cell death (White et al., 1979). Thus, measurement of the lipid biomarkers, such as PLFAs, provides quantitative measures of the viable biomass as well as information on community composition and physiological status of the microbiota although this approach is limited by its low resolution and demanding technical procedure (Guckert et al., 1985; Pinkert et al., 2002; White et al., 1997). Another limitation of most intestinal microbiota research is that fecal biota was used as a surrogate for the entire gut microflora. However, mucosa-associated microbial populations that are in close contact with the underlying gut epithelium could be distinct from luminal populations and may fulfill different but significant roles within the gut ecosystem (Eckburg et al., 2005; Scupham et al., 2006). Thus, in this study, we chose whole intestinal sections including both mucosal tissues and luminal contents for PLFA analysis and elucidated novel information regarding viable microbiota in the chicken intestine in response to tylosin treatment. First, long-term use of tylosin did not significantly change the total bacterial biomass in jejunum and ileum (Fig. 1A), which argues one of hypotheses that AGP use may reduce the maintenance cost of gastrointestinal tract system via reduced total bacterial load in intestine (Collier et al., 2003; Gaskins et al., 2002). We speculate that the use of AGP may temporarily reduce bacterial load but total bacterial populations eventually rise to normal level with a different microbial structure. Second, PLFA analysis revealed similar bacterial biomass among different intestinal locations. A recent study using plate counts (Dumonceaux et al., 2006) showed viable counts increased one to three orders of magnitude along the chicken gastrointestinal tract from the proximal (e.g., jejunum) to distal small intestine (e.g., ileum) and further increased one to two orders of magnitude from the distal ileum to the cecum. This discrepancy is likely due to the selectivity and insensitivity of cultivation and the lack of evaluation of mucosa-associated bacterial populations in previous study (Dumonceaux et al., 2006). Third, interestingly, PLFA analysis revealed abundant fungal biomass in the chicken intestine regardless of AGP usage (Fig. 1). Unlike bacterial flora, characterization of the fungal ecosystem in the intestine has been poorly studied. The significance of intestinal fungi in host health and growth performance is still not well understood. Scupham et al. (2006) recently provided the first culture-independent analysis of fungal rDNA from mammalian intestine, showing that diverse fungal taxa are present in the murine intestine with significantly higher abundance in the mucus biofilm than that in luminal feces. This finding and our PLFA analysis strongly suggest that fungi may play significant roles in enteric microbial functions. It has been demonstrated that anaerobic fungi contribute to the fiber digestion in ruminants (Weimer, 1998). Thus, fungi may also play a role in food metabolism for nutrients and energy bioavailability in nonruminant animals. The effect of AGPs on fungal diversity and relative abundance needs to be examined in future studies.
It is not surprising that the lactobacilli population was reduced in AGP-treated chickens based on the16S rDNA analysis used in this study. Previous independent chicken studies also showed that AGP usage significantly reduced the population of Lactobacillus species (Danzeisen et al., 2011; Dumonceaux et al., 2006; Engberg et al., 2000; Guban et al., 2006; Knarreborg et al., 2002; Zhou et al., 2007). It has been well documented that the Lactobacillus are the primary commensal bacteria for production of bile salt hydrolase (BSH) (Begley et al., 2006; Gaskins et al., 2002; Ridlon et al., 2006). The conjugated bile salts, which are required for efficient lipid emulsification and utilization, could be catalyzed by BSH and transformed to unconjugated bile salts that are much less effective for lipid metabolism (Begley et al., 2006; Gaskins et al., 2002; Ridlon et al., 2006). Thus, the conjugated bile salts are needed to maintain efficient lipid digestion and the absorption of fatty acids. Therefore, the decreased lactobacilli population in AGP-treated animals likely reduce intestinal BSH activity, which would increase the relative abundance of conjugated bile salts, consequently promoting lipid metabolism and energy harvest, and enhancing body weight gain of food animals. In agreement with this hypothesis, previous animal studies (Dumonceaux et al., 2006; Engberg et al., 2000; Guban et al., 2006; Knarreborg et al., 2002; Zhou et al., 2007) have consistently demonstrated that growth promoting effect of AGPs was highly correlated with decreased activity of intestinal BSH activity. In light of the findings from this pilot study and the published information, we speculate that BSH inhibitors are promising alternatives to AGPs for mitigating antimicrobial resistance in food animals. This hypothesis needs to be examined in the future.
Despite the novel and meaningful findings from this pilot study, the use of a small sampling number and sequencing of a relatively low number of clones from 16S rDNA library could not provide an in-depth view of the gut microbial ecology. To overcome this limitation and confirm the findings from this study, the recently developed high-throughput approaches, such as 16S rDNA taq pyrosequencing (Acosta-Martinez et al., 2008), metagenomic sequencing of gut microbiota (Turnbaugh et al., 2006), and GeoChip (He et al., 2010), will facilitate us to accurately elucidate phylogenetic, functional, and metabolic changes of gut microbiota affected by AGP usage. The information obtained from future large-scale experiments using the cutting-edge modern molecular approaches and the growth-relevant intestinal samples will guide us to develop effective alternatives to AGPs for enhancing the safety and sustainability of food animal.
Footnotes
Acknowledgments
We are grateful to Dr. David C. White (Deceased, Center for Biomarker Analysis, The University of Tennessee) for his stimulating discussion and collaboration on this project. We thank Aaron Peacock and Amanda Smithgall for technical assistance for PLFA analysis. This study was supported by University of Tennessee Professional Development Award and UT Obesity and Research Center Pilot Grant.
Disclosure Statement
No competing financial interests exist.