
Abstract
The presence of Staphylococcus aureus in raw milk can represent a potential threat to human health, due to the introduction of pathogenic strains into dairy food supply chain. The present study was performed to investigate the genetic variation among S. aureus strains isolated from bulk tank goat's milk. The virulence profiles were also assessed to link the isolates with the potential source of milk contamination. A population study was performed on 60 strains using distance-based methods such as pulsed-field gel electrophoresis (PFGE), and the output was analyzed using Structure statistical software (University of Chicago;
Introduction
S
The prevalence of S. aureus in bulk tank goat's milk ranges between 32% (Muehlherr et al., 2003) and 98.8% (Jackobsen et al., 2011). S. aureus produces a large array of virulence factors such as enterotoxins, hemolysins, and leukocidins, which play an important role in determining human disease. Milk storage at refrigeration temperatures is a critical aspect in controlling S. aureus multiplication and preventing the production of enterotoxins (Jørgensen et al., 2005). The pathogenicity profile of S. aureus can be host-specific (van Leeuwen et al., 2005); therefore, genes coding for toxin production, combined with accessory gene regulator (agr) alleles, can be considered important determinants to define biovars based on the most likely origin of the strains (Larsen et al., 2000; Bhatia and Zahoor, 2007; Spanu et al., 2012). Literature on the characterization of S. aureus virulence factors is available on strains isolated from mastitic dairy ruminants (Alves et al., 2009; Ote et al., 2011; Virdis et al., 2010); little research is focused on isolates recovered from bulk tank goat's milk.
Pulsed-field gel electrophoresis (PFGE) is the gold standard method for molecular subtyping of foodborne bacteria (Hyytiä-Trees et al., 2007). Other tools, such as Bayesian clustering models, implemented by Structure statistical software (University of Chicago;
Materials and Methods
Staphylococcus aureus isolation
Twenty-six goat farms located in Sardinia, Italy, were visited monthly for 3 consecutive months to collect bulk tank milk samples. During each visit, a single bulk tank milk sample (200 mL) was collected, transported refrigerated to the laboratory, and analyzed for the detection and enumeration of Coagulase Positive Staphylococci (CPS) according to ISO 6888-1/2 (1999a,b) within 6 h after collection. From each positive sample, five CPS colonies were selected, subcultured on brain-heart infusion (BHI) agar (Oxoid, Basingstoke, UK), and identified by standard microbiological procedures. Each determined isolate was subjected to thermolabile and thermostable DNase production testing (Bio-Rad Laboratories, Hercules, CA) for free (bioMérieux, Lyon, France) and bound (Oxoid, Basingstoke, UK) coagulase reactions. The strains were also identified by biochemical test API ID32 STAPH (bioMérieux) and tested for the presence of Staphylococcus aureus gyrA gene by polymerase chain reaction (PCR) (Jarraud et al., 2002). After identification, strains were stored at −80°C in BHI broth with glycerol (15% v/v). In order to avoid overrepresentation, a single random strain from each positive sample was selected for further analysis.
PFGE
Cluster analysis was conducted by PFGE according to the harmonization protocol suggested by Murchan et al. (2003) with minor modifications; total genomic DNA was digested using smaI restriction enzyme (Invitrogen Ltd., Paisley, UK) for 3 h at 25°C. Plugs were loaded into 0.8% agarose gel (Bio-Rad Laboratories, Hercules, CA), where the run performed in Tris Borate EDTA (TBE) 0.5×, and S. aureus NCTC 8325 was used as a reference standard strain. Electrophoresis run was performed using CHEF Mapper XA System (Bio-Rad Laboratories), and the DNA banding patterns were analyzed with Gel Compar II software 6.5 (Applied Math, Kortrijk, Belgium). Statistically, the Pearson's correlation coefficient was calculated with 1.0% of optimization setting, and represented by Unweighted Pair Group Method using Arithmetic averages (UPGMA). Cut-off of 70% similarity was used to obtain PFGE clusters. Isolates with identical restriction profile were assigned to the same pulsotype (PT). The difference of at least one band was used as the criterion to distinguish a pulsotype from another. PFGE profiles were also computed and visually compared to categorize the isolates as indistinguishable, closely related, possibility of related, or unrelated (Tenover et al., 1995).
Population structure and genetic diversity analysis
Visual examination of the DNA restriction patterns generated by PFGE was carried out in order to build a binary matrix used to infer population structure based on genetic information. The presence of a band was coded as “1” and its absence as “0.” Bayesian clustering model implemented by Structure 2.3.3 software (Pritchard et al., 2000) was used to investigate the genetic population structure. In the current study, the word “population” is intended to describe a group of related microorganisms, such as bacteria of the same species. The admixture model was performed to estimate the number of genetically homogeneous cluster (K), without using a priori information on the cluster numbers of the population. The model was run with 10 replications of K ranging from 1 to 15. For each run, 300,000 MCMC interactions and a burn-in period at 30,000 were set. The K value with the highest likelihood was determined according to the method described by Evanno et al. (2005). The genetic divergence between groups inferred by Structure was estimated by analysis of molecular variance (AMOVA) with Arlequin software 3.5 (Excoffier and Lischer, 2010). The genetic differentiation between and within groups was evaluated as the fixation index (F ST).
Virulence factors
Total genomic DNA was extracted according to the protocol proposed by De Buyser et al. (1989).
Different multiplex PCR were prepared, in order to determine the agr alleles and the virulence factors genes of S. aureus isolates. Primers used to detect genes coding for Agr (agr I–IV), SEs (sea-see), SEs-like (seh, sek, sel, sem, seo, sep) and other virulence factors such as leukocidins (pvl, lukE/D, lukM), hemolysins (hlb, hlg, hlgv), exfoliatins (eta, etb, etd), epidermal cell differentiation inhibitor genes (edinABC), and toxic shock syndrome toxin-1 (tst) were previously described (Jarraud et al., 2002; Wu et al., 2011). PCR reagent concentrations were as described by Kérouanton et al. (2007). DNA amplification was performed using a Thermal Cycler Gene Amp PCR 9700 (Applied Biosystems, Carlsbad, CA) under the following conditions: initial denaturation at 94°C for 5 min followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, extension at 72°C for 1 min, and a final extension at 72°C for 7 min. PCR products were resolved on 1.5% agarose gel, visualized by ultraviolet transilluminator (Gel DOC XR), and analyzed using software Quantity One (Bio-Rad Laboratories). The presence of a given amplicon was coded as “1” and its absence as “0”. The linear combination of the binary results for each gene allowed to obtain the pathogenicity profile of the isolates. Moreover, the relationship between genes associated with a specific host (Fueyo et al., 2001; Bania et al., 2006) was used to describe four different biovars. The presence of sec and/or sel genes was used to define the “animal enterotoxigenic” biovar; lukE/D, lukM, hlb, hlgv genes the “animal non enterotoxigenic” biovar; sea, sem, seo, hlg genes the “human” biovar; and “non-host specific” biovar were the strains sharing hlb, hlg, hlgv genes (Spanu et al., 2012).
Detecton of Staphylococcal enterotoxins
In vitro production of Staphylococcal enterotoxins (SEs) A–E was determined on BHI broth cultures by ELISA immune-enzymatic kit (Ridascreen Set; R-Biopharm, Darmstadt, Germany) according to the manufacturer's instructions. Briefly, S. aureus broth cultures were incubated at 37°C for 18–24 h. After centrifugation, the supernatant was filtered on nitrocellulose membranes (0.22 μm) and tested for the presence of the SEA-E. Optical density were measured at 450 nm with Sunrise Plates Reader (TECAN, Grödig, Austria), and enzyme immunoassays were evaluated using Ridasoft Win software (R-Biopharm).
Detection of MRSA
The isolates were tested for the presence of the gene encoding methicillin resistance (mecA) using a simplex PCR (Vandenesh et al., 2003). Minimal inhibitory concentrations (MICs) of oxacillin (Sigma-Aldrich, St. Louis, MO) were also determined by broth microdilution method according to Clinical Laboratory Standards Institute guidelines (CLSI, 2006, 2007). Reference strains S. aureus American Type Culture Collection (ATCC) 29213 and Enterococcus faecalis ATCC 29212 were used as quality controls.
Results
Staphylococcus aureus isolation
The presence of CPS was observed in 60 (76.9%) out of 78 milk samples collected from 26 different dairy goat farms. CPS count was 2.9±1.1 (
Population study
Using PFGE analysis, five different clusters (CLs) were identified. The most represented was CL1, consisting of 31 strains (52.5%), followed by CL5 with 23 strains (38.3%). The remaining three clusters included one to three isolates (Fig. 1). Comparing the DNA banding pattern, a wide genetic diversity of the strains was observed. The isolates were divided into 25 different pulsotypes, where the most represented pulsotype was the PT2, with 16 strains grouped in two different CLs. Overall pulsotypes were not associated with the farm of origin. The number of pulsotypes within each CL is reported in Figure 1. According to the criteria proposed by Tenover et al. (1995), most of the strains (93.3%) could be classified as closely related, while four (6.6%) were considered possible related. The matrix obtained by PFGE band scoring consisted in 46 markers among 60 strains. Population structure analysis showed a trend in the maximum likelihood at K=2, corresponding to ΔK=86.26. Two distinct genetic groups were identified, the first (GRP A) consisted of 34 strains, while the second (GRP B) included 26 strains. The estimated average coefficient of membership (Q) was 0.95±0.07 (
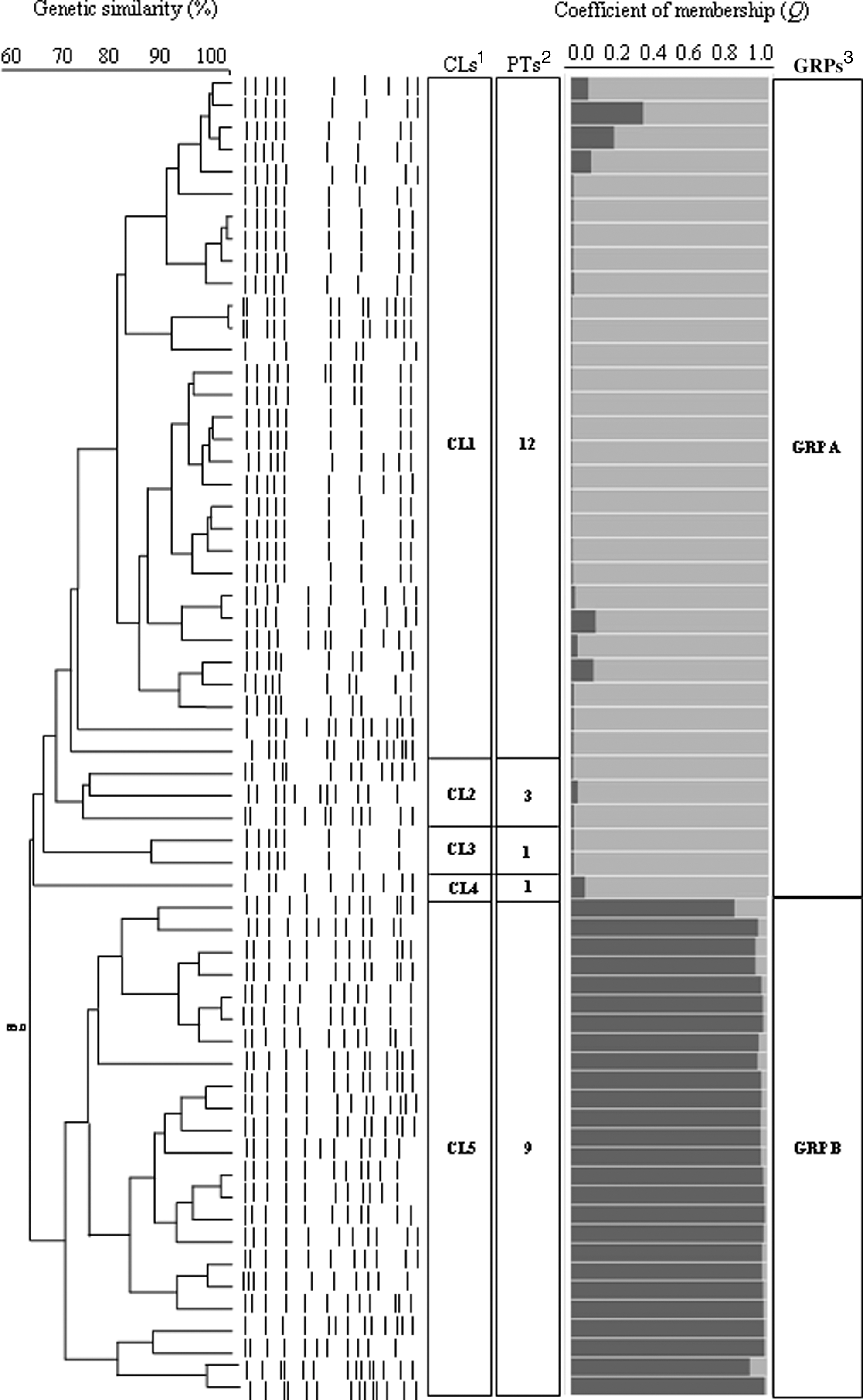
Unweighted Pair Group Method with Arithmetic Mean (UPGMA) clustering and population structure analysis of 60 Staphylococcus aureus strains isolated from bulk tank goat's milk. 1CLs, pulsed-field gel electrophoresis clusters; 2PTs, pulsotypes within each cluster; 3GRPs, groups obtained by analysis with Structure statistical software (University of Chicago;
Fixation index (F ST), 0.29393.
Virulence profile and biovars
The binary matrixes produced by the presence or absence of the 23 investigated genes allowed us to obtain 18 different pathogenicity profiles. Genes coding for SEs B, E, and M, Panton Valentine Leukocidin, and exofoliatins A, B, and D were never detected. The frequency distribution for each of the virulence factors detected stratified by agr locus is summarized in Table 2. On the basis of the selected genes, 45 (75%) strains belonged to the “animal biovar,” whereas 25 (41.7%) of them were ascribed to “animal enterotoxigenic” and 20 (33.3%) to the “animal non-enterotoxigenic” biovars. The “non-host specific” biovar was represented by 11 (18.3%) strains, while only 4 (6.7%) strains could be included in the “human” biovar.
agr locus, accessory gene regulator locus.
Detection of Staphylococcal enterotoxins
Immune-enzymatic detection of SEA-SEE showed that 23 S. aureus strains (38.3%) were SEC producers, three (5.0%) were SEA producers, and only one (1.7%) was positive for SED production. All strains producing SEs carried the corresponding SE genes. Enterotoxins B and E were never detected.
Susceptibility to oxacillin and mecA encoding gene
All the tested strains were shown to be oxacillin-sensitive, with values of MIC50, MIC90, mode equal to 0.25 μg/mL and range of the MICs ranging from 0.12 to 0.25 μg/mL. None of the isolates carried the mecA gene encoding methicillin resistance.
Discussion
The prevalence of S. aureus strains in bulk tank goat's milk samples observed in the present study was 76.9%, which is in the range previously reported (Muehlherr et al., 2003; Scherrer et al., 2004; Jackobsen et al., 2011). Therefore, goat's milk may represent a potential source of S. aureus introduction into the dairy supply chain. PFGE is a powerful discriminatory method for subtyping microorganisms and evaluating interstrain genetic relationship (Mørk et al., 2005). The selected 60 S. aureus strains of the present study were ascribed to 25 different pulsotypes, indicating wide variations among strains. The differences in the banding pattern ranged from one to six bands, indicating the possible occurrence of a single or two independent genetic events (Tenover et al., 1995). Despite this variability, strains were grouped into two main clusters and categorized as closely or possible related, suggesting that the isolates could be derived from a common ancestor. A complementary approach to study the structure of a population is the Bayesian clustering model provided by Structure statistical software, which assigns individuals into groups, conferring a coefficient of membership. The model run with PFGE data estimated the presence of two distinct groups among S. aureus isolates. The computed mean coefficient of membership was high (Q=0.95), providing evidence of two well-defined genetic groups. Using a cut-off of Q=0.88, no admixed individuals were observed, with the exception of one strain with Q=0.63. Interestingly, using a similarity coefficient of 65% to build the dendrogram, two main clusters could be defined, which perfectly overlap with the partition estimated by the Bayesian clustering model. In order to explore the genetic diversity between groups, AMOVA was performed. AMOVA analysis indicates that, although two distinct groups can be defined, a great part of genetic variability was attributable to the intra-group source of variation (70%). This is in agreement with the great sub-partition showed by the dendrogram within each cluster. The relationship between virulence pattern and host specificity is well documented (Larsen et al., 2000; Hennekinne et al., 2003; Bhatia and Zahoor, 2007), and it can be used to predict the most likely source of milk contamination (Spanu et al., 2012). Most of the strains (75%) could be reconducted to the “animal” biovar. Among these, the strains harboring genes encoding enterotoxins (“animal enterotoxigenic”) accounted for 55.5%. As expected, the prevalence of “human” biovar strains was low (6.7%). In addition, the high prevalence of S. aureus strains belonging to the agrIII and agrI alleles (93.3%), which is characteristic of isolates from small ruminants (De Santis et al., 2005; Vautor et al., 2007, 2009), confirms that the main source of S. aureus in milk is accountable to contamination of animal origin. About 45.0% of the strains showed in vitro enterotoxins production, which may pose a concern to human health, as a consequence of the introduction of potentially harmful strains into dairy supply chain. Therefore, the high prevalence of enterotoxigenic S. aureus strains detected in bulk tank goat's milk should focus the attention of the goat's milk industry on promoting the implementation, at farm level, of best management practices. Additional information on the pathogenicity of S. aureus isolates was obtained by investigating the presence of mecA gene and associated resistance to oxacillin. All selected strains were susceptible to methicillin, which confirms previous data showing that the mecA gene is mainly associated with S. aureus isolated from humans (Vandenesh et al., 2003), while it is rarely recovered from dairy small ruminants (Virgin et al., 2009; Virdis et al., 2010).
Conclusion
The study showed the presence of two distinct genetic clusters among S. aureus strains isolated from bulk tank goat's milk. The combination of PFGE with Structure statistical software proved to be a useful tool to investigate genetic variability among strains. Based on the virulence profile, most of the strains recovered were of animal origin, confirming the existence of a specific host-pathogen relationship in dairy goats. The presence of enterotoxin producer strains raises concern about the potential impact of goat's milk on human health. The use of other typing methods, such as spa yyping and multi locus sequence typing, integrated with Structure statistical software analysis could be an interesting prospective for further investigations aimed to conduct extensive phylogenetic studies.
Footnotes
Acknowledgments
This work was funded by Fondanzione Banco di Sardegna 2010 and Programma Operativo FSE Sardegna 2007–2013, Legge Regionale, 7 Agosto 2007 (No. 7, Research Progress and Technological Innovation in Sardinia).
Disclosure Statement
None of the authors of this paper has a financial or personal relationship with other people or organizations that could inappropriately influence or bias the content of the paper.