
Abstract
The role of capsular polysaccharides and lipooligosaccharides in cell surface hydrophobicity, surface charge, autoagglutination (AAG), and attachment to abiotic surfaces of three strains of Campylobacter jejuni and one strain of C. coli were investigated. This was achieved by removal of capsular polysaccharides and truncation of lipooligosaccharides core oligosaccharides by inactivation of the kpsE and waaF genes, respectively. The mutants and the wild-type strains were compared after growth under planktonic (broth) and sessile (agar) conditions. Cells grown as planktonic cultures showed a significantly (p<0.05) higher degree of hydrophobicity and AAG activity but differed from their sessile counterparts with respect to surface charge and attachment counts, depending on the strain. These results suggest that prior mode of growth affects the surface properties and attachment of Campylobacter in a strain-dependent manner. There were no significant (p>0.05) differences between the three C. jejuni strains and their ΔkpsE and ΔwaaF mutants with respect to all traits tested. Inactivation of the kpsE gene significantly (p<0.05) reduced the surface charge of the C. coli strain from ∼-10 to ∼-6 mV and increased its AAG activity, while disruption of the waaF gene significantly (p<0.05) increased its surface hydrophobicity by >8° and decreased the numbers of cells attaching to stainless steel and glass by ∼0.5 log/cm2. These results suggest that surface polysaccharides may influence the surface properties and attachment to abiotic surfaces of C. coli but not C. jejuni. This suggestion, however, requires further investigation using a larger number of strains of both species.
Introduction
C
Capsular polysaccharides (CPS) and lipooligosaccharides (LOS) are found to have an impact on cell surface hydrophobicity, AAG, and attachment of many Campylobacter strains to cell lines. For example, inactivation of CPS expression by mutation of the CPS transport genes kpsM, kpsS, and kpsE resulted in an increased surface hydrophobicity and reduced ability of C. jejuni strains 81–176 and 11168 to attach to epithelial cells INT 407 (Bacon et al., 2001; Bachtiar et al., 2007; Jeon et al., 2009). Truncation of LOS core oligosaccharides of these strains by inactivation of the heptosyltransferase II gene waaF also resulted in a similar effect (Jeon et al., 2009; Kanipes et al., 2004). In addition, removal of CPS and sialic acid from LOS by inactivation of the genes kpsM and neuC, respectively, reduced AAG activity of C. jejuni 81–176 (Guerry et al., 2006). However, the role of CPS and LOS in attachment of C. jejuni and C. coli to abiotic surfaces has not been investigated.
We have recently reported that attachment of different Campylobacter strains to abiotic surfaces, such as stainless steel and glass, is associated with increased cell surface hydrophobicity (Nguyen et al., 2010; Nguyen et al., 2011). The highly hydrophobic and adherent strains also attached strongly to the agar surface when they were grown on modified charcoal cephoperazone deoxycholate agar–Preston plates (Nguyen et al., 2011). Since surface polysaccharides including CPS and LOS are associated with cell surface hydrophobicity (van Oss, 1995) and the adherent nature of Campylobacter (Guerry and Szymanski, 2008), we undertook the present study to investigate whether CPS and LOS have a role in affecting Campylobacter surface hydrophobicity, surface charge, AAG, and mediating attachment to two abiotic surfaces, namely, stainless steel and glass. This was achieved by inactivation of CPS expression and truncation of LOS core oligosaccharides by disruption of the kpsE and waaF genes, respectively, in four highly hydrophobic and adherent Campylobacter strains (three C. jejuni strains and one C. coli strain).
Materials and Methods
Bacterial strains and culture conditions
The bacterial strains and plasmids used in this study are listed in Table 1. For all assays, Campylobacter strains were grown as planktonic cultures in Nutrient Broth No. 2 (NB2, Oxoid) (without shaking) or as sessile cultures on NB2 supplemented with 1.2% (wt/wt) technical agar (Agar No. 3, Oxoid) at 42°C for 48 h under capnophilic conditions (5% CO2). Cells were harvested by centrifugation (13,131×g, 10 min), washed once with 150 mM phosphate buffered saline (PBS, 8.5 g/L NaCl, 0.2 g/L KCl, 1.44 g/L Na2HPO4, 0.24 g/L KH2PO4, pH 7.4) and then resuspended in the same buffer unless otherwise stated. Escherichia coli strains were grown in Luria-Bertani (Sigma, St. Louis, MO) broth with shaking (150 rpm) or on Luria-Bertani agar at 37°C for cloning experiments. Media were supplemented with ampicillin (100 μg/mL) or kanamycin (50 μg/mL) where appropriate.
Penner serotyping based on soluble heat-stable (HS) antigens.
Kan, kanamycin; Amp, ampicillin.
Construction of ΔkpsE and ΔwaaF mutants
Construction of ΔkpsE mutants was performed as previously described (Bachtiar et al., 2007). A suicide plasmid pGEM-T::kpsE::KmR was constructed by cloning a 614-bp upstream DNA fragment (including 400 bp of the 5′ end of the kpsE gene and 214 bp of the 3′ end of the flanking kpsT gene), the kanamycin cassette (from the pMW2 vector), and a 348-bp downstream fragment (containing 251 bp of the 3′ end of the kpsE gene and 97 bp of the 5′ end of the flanking kpsD gene) into pGEM-T Easy vector (Promega) (Table 2). The suicide plasmid was subsequently introduced to Campylobacter cells by electroporation as previously described (van Vliet et al., 1998) to replace a 462-bp segment of the kpsE gene by a 1448-bp kanamycin resistance gene.
The genome of Campylobacter jejuni NCTC 11828 (Pearson et al., 2007) was used to design all of the primers except for waaF_F2 and waaF flank_R, which were designed using the genome of C. jejuni ICDCCJ07001 (Zhang et al., 2010),
Restriction sites are underlined.
Truncation of LOS core oligosaccharides of C. jejuni strains was achieved by inactivation of the heptosyltransferase II gene (waaF) using a similar protocol to that described above for mutation of the kpsE gene.
Detection of CPS and LOS by Alcian Blue staining
The differences between CPS and LOS expression by the wild-type strains and the ΔkpsE and ΔwaaF mutants were determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by Alcian Blue staining as described previously (Karlyshev and Wren, 2001). Briefly, bacterial cells from 48-h planktonic and sessile cultures were resuspended in lysis buffer (31.25 mM Tris-HCl [pH 6.8], 4% SDS, 20% glycerol, and 0.025% bromophenol blue). Samples were heated at 100°C for 5 min and centrifuged at 13, 131×g for 5 min. Aliquots of 20 μL of the supernatant were taken and mixed with 1 μL of proteinase K (20 mg/mL, Sigma, St. Louis, MO). The samples were incubated at 50°C for 1 h and fractionated on precast SDS-PAGE (Bio-Rad, Criterion, 4–20%% Tris-HCl, Cat. No. 3450033) followed by staining with 0.1% Alcian Blue (Sigma) in 40% ethanol and 5% acetic acid.
Cell surface hydrophobicity
Cell surface hydrophobicity of Campylobacter was determined using water contact angle measurement as described by Busscher et al. (2008) using a goniometer (KSV Instruments, Ltd., Helsinki, Finland). Planktonic and sessile cells grown as described above were resuspended in ultrapure (type I) Milli-Q water and filtered through a 0.45-μm-pore membrane filter (mixed cellulose ester HA filter, 25 mm diameter) (Millipore, Billerica, MA). The bacterial cell-filters were attached to glass using double-sided adhesive tape and dried in a desiccator containing drierite (CaSO4, W. A. Hammond Drierite Company Ltd., Xenia, OH) for 30 min. A drop of Milli-Q water was placed onto the filter, and the contact angle was determined by measuring the angle between the filter surface and the tangent to the drop at the solid–liquid air point.
Cell surface charge
Cell surface charge of Campylobacter was determined using zeta potential measurement as previously described (Li and McLandsborough, 1999). A volume of 1 mL of cell suspension (107 colony-forming units/mL in PBS) was used to measure zeta potential using a Zetasizer Nano (Malvern Instruments Ltd., Worcestershire, UK).
AAG activity
AAG activity was determined using an assay described by Misawa and Blaser (2000). Planktonic and sessile cells were resuspended in 150 mM PBS (pH 7.4) to an OD600 1.0. Two milliliters of cell suspension was transferred to 12×75-mm polypropylene tubes, and incubated at 25°C for 0.33, 2, 4, 6, and 24 h. After each incubation time, 1 mL of the upper phase was carefully removed and the OD600 was measured. The reduction of the OD600 in the upper phase from the initial setting of 1.0 reflects the degree of AAG activity.
Bacterial attachment to abiotic surfaces
The ability of Campylobacter to attach to abiotic surfaces was determined based on the numbers of cells attaching to stainless steel (type 304, #4 finish, 50×20×0.9-mm thickness) (Hanly Sheet Metal, Murrarie, Queensland, Australia) and glass (76×25×1-mm thickness) (Sail Brand, China) as previously described (Nguyen et al., 2011).
Attachment assays were performed by placing the clean sterile surfaces in the vertical position in 40 mL of cell suspensions (107 colony-forming units/mL) in PBS contained in 50-mL centrifugation tubes (Sarstedt, Nümbrecht, Germany) at 25°C for 20 min. After attachment, the coupons were removed using sterile forceps and rinsed twice by gently dipping into 40 mL PBS to remove excess liquid droplets and loosely attached cells. The coupons were subsequently stained for 20 min in the dark using 0.01% (wt/vol) acridine orange stain, followed by gently dipping the coupons in 40 mL of Milli-Q water. The attached cells were quantified using epifluorescence microscopy (Leica DM 6000B, Leica Microsystems CMS GmbH, Wetzlar, Germany). A total of 50 fields per coupon were scanned and the fluorescent cells were enumerated. Counts were calculated as log cell/cm2. The level of detection is 2.05 log cell/cm2, which is obtained when the total number of attached cells in 50 microscopic fields is one.
Statistical analysis
All experiments were performed in triplicate. All quantitative data, unless otherwise stated, are presented as means with error represented by standard deviation. Significant differences between all data sets were determined by means of one-way analysis of variance (Tukey's method) using Minitab software (Minitab 15; Minitab Inc., Minneapolis, MN).
Results
Construction of ΔkpsE mutants and ΔwaaF mutants
Mutants (ΔkpsE) of all four Campylobacter strains (C516a, 81116, C977a, and C719a) were successfully generated. As a consequence of insertion of the kanamycin cassette in the kpsE coding region, the ΔkpsE mutants generate a 2.4-kbp polymerase chain reaction (PCR) product using primers kpsT_F and kpsD_R as opposed to a 1.4-kbp PCR products generated by the wild-type strains (Fig. 1A).
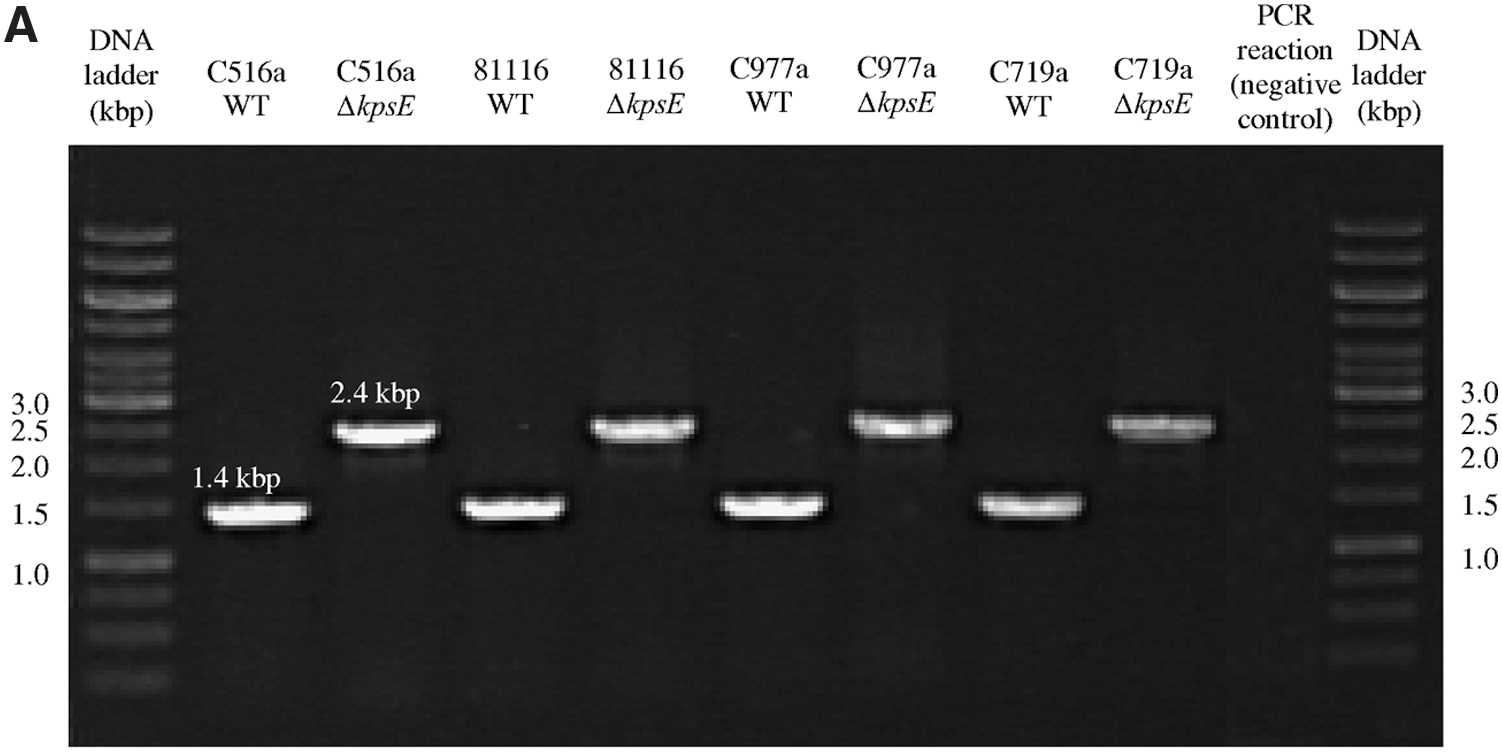
Polymerase chain reaction (PCR) verification of the insertion of the kanamycin cassette in the ΔkpsE and ΔwaaF mutants. The ΔkpsE mutants showed 2.4-kbp PCR products using primers kpsT_F and kpsD_R as opposed to 1.4-kbp PCR products shown by the wild-type (WT) strains
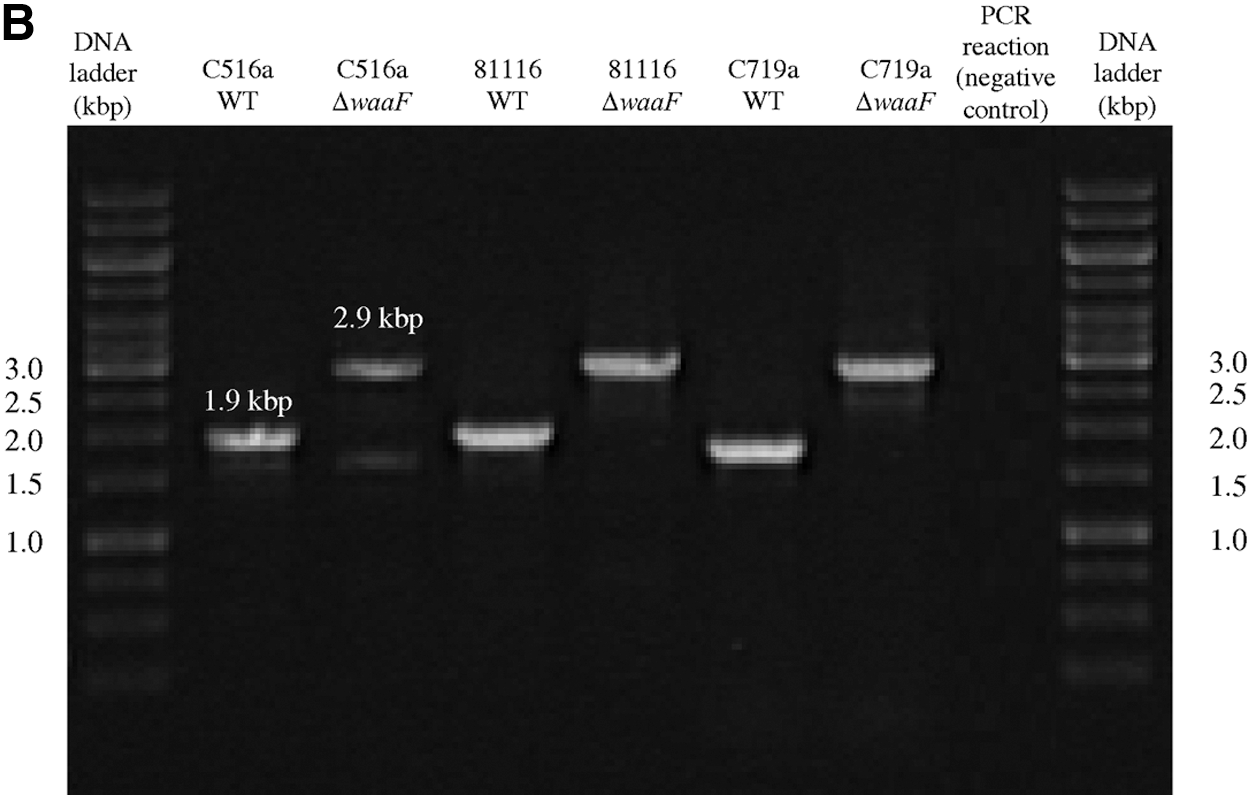
Mutants (ΔwaaF) of the two C. jejuni strains (C516a and 81116) and the C. coli strain (C719a) were also successfully generated using a homologous double crossover event between the plasmids pGEM-T::waaF1::KmR (for C. jejuni) and pGEM-T::waaF2::KmR (for C. coli). As a consequence of homologous DNA recombination, a 503-bp fragment of the waaF gene in the two C. jejuni strains and a 286-bp fragment of the waaF gene in the C. coli strain were deleted and replaced by a 1448-bp kanamycin resistance gene. As a consequence of insertion of the kanamycin cassette in the waaF coding region, the ΔwaaF mutants showed a 2.9-kbp PCR product using primers waaF_F and gmhA_R (for C. jejuni) and primers waaF_F and waaFflank_R (for C. coli) as opposed to a 1.9-kbp PCR products shown by the wild-type strains (Fig. 1B). Several attempts to generate a ΔwaaF mutant for C. jejuni C977a were unsuccessful because no pGEM-T::waaF1::KmR transformants could be isolated.
Inactivation of the kpsE and waaF genes results in loss of CPS and truncation of LOS core oligosaccharides
CPS and LOS expression by the wild-type strains, ΔkpsE and ΔwaaF mutants grown as sessile cultures, was assessed by SDS-PAGE followed by Alcian Blue staining (Fig. 2). Using this technique, the CPS of all the wild-type strains appeared as high-molecular-weight smears (for example, the smear running above 35 kDa [for C977a] and above 25 kDa [for C719a]). The LOS appeared as low-molecular-weight (below 10 kDa) smears. The bands above 10 kDa may be aggregates of LOS due to overloading of the gel, but may alternatively be CPS molecules, which also run between 10 and 35 kDa. As shown in Fig. 2, the mutation of the waaF gene had an effect on all three of the strains (C516a, 81116, and C719a) as indicted by the disappearance of the bands above 10 kDa. However, it is difficult to discern what effect the kpsE mutation had on all of the strains. This is because mutation of the kpsE gene had no observable effect on the CPS and LOS profiles of the C516a and 81116 strains and minimal effect on the C977a and C719a strains. These features are shown as smears disappearing above the 35-kDa region and around the 35-kDa region, respectively. When grown in planktonic cultures, cells of all strains showed comparable CPS and LOS profiles as the sessile cells (data not shown).
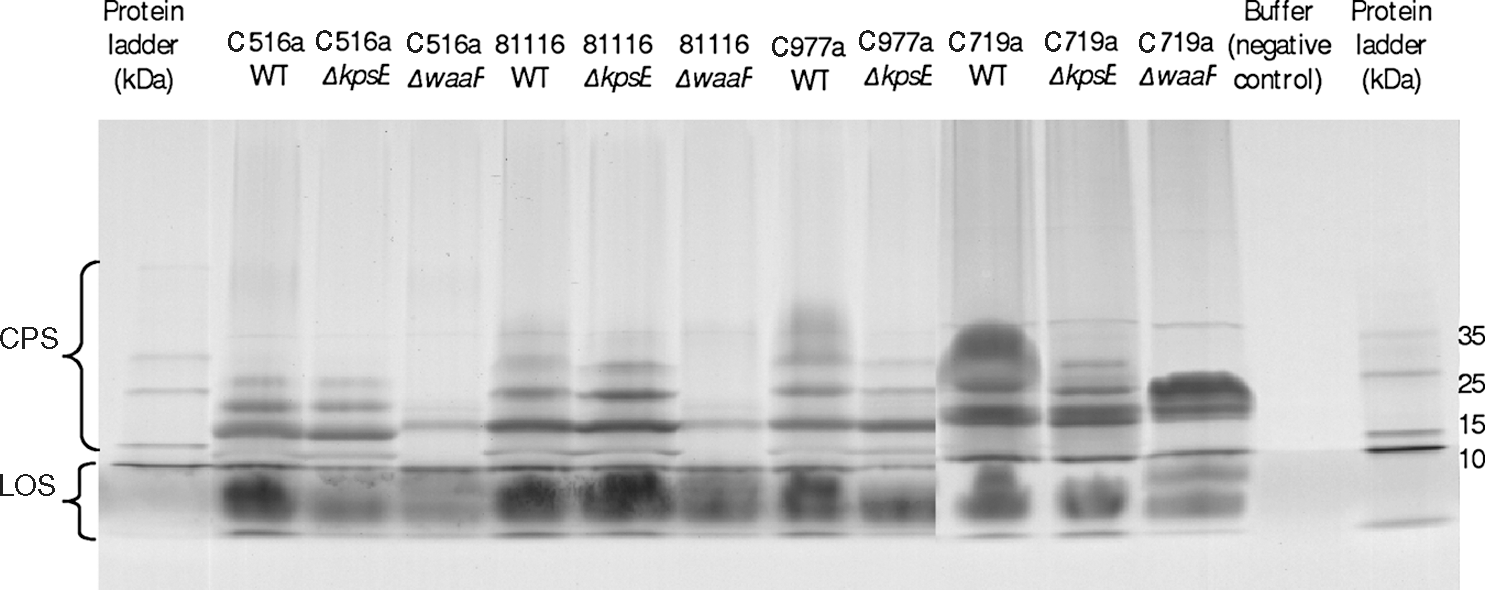
Effects of the insertional mutation in the kpsE and waaF genes on the expression of capsular polysaccharides (CPS) and lipooligosaccharides (LOS) by Campylobacter. The CPS and LOS were fractionated on sodium dodecylsulfate–polyacrylamide gel electrophoresis and visualized using Alcian blue staining. The first and last lanes are protein markers. The second last lane is a negative control. The remaining lanes are wild-type and deletion mutant CPS and LOS fractions of the Campylobacter strains and labeled by strain number.
Inactivation of the kpsE and waaF genes affect the cell surface properties, AAG, and attachment to abiotic surfaces for the C. coli strain but not C. jejuni strains
In order to determine the role of CPS and LOS on cell surface properties, AAG, and attachment of Campylobacter to abiotic surfaces, water contact angle, zeta potential values, AAG activity, and counts of attached cells to stainless steel and glass of the wild-type strains, ΔkpsE and ΔwaaF mutants were compared. As shown in Table 3 and Fig. 3, there were no significant (p>0.05) differences in all traits tested between the wild-type, ΔkpsE, ΔwaaF mutants of three C. jejuni strains grown in either planktonic or sessile culture, indicating that inactivating the kpsE and waaF genes did not affect surface hydrophobicity, surface charge, and attachment of C. jejuni to abiotic surfaces. Inactivating the kpsE gene also did not affect surface hydrophobicity and attachment of the C. coli strain but reduced the negative charge on the cell surface (from ∼-10 mV to ∼-6 mV) and increased AAG activity (from OD600 of ∼0.924 to ∼0.204). In contrast, inactivating the waaF genes did not affect cell surface charge and AAG of the C. coli strain but increased its surface hydrophobicity by >8° and reduced its ability to attach to abiotic surfaces by ∼0.5 log/cm2.
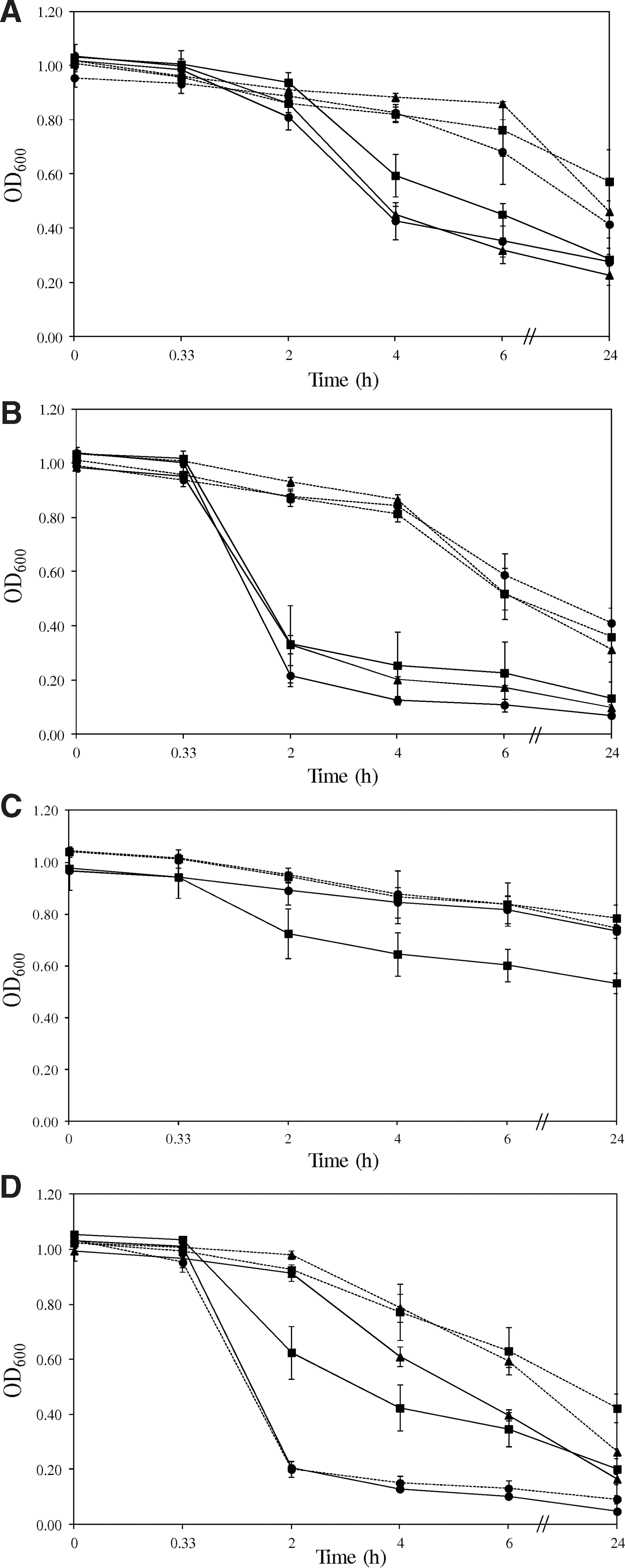
Autoagglutination activity of the wild type (▪), ΔkpsE (•) and ΔwaaF (▴) mutants grown as planktonic (solid lines) and sessile (broken lines) cultures for Campylobacter jejuni C516a
Data labeled with the same letter are not significantly different (p>0.05). The comparison included the entire set of strains and both planktonic and sessile modes of growth.
Cell surface hydrophobicity determined using water contact angle measurement (wCAM). Cells were grown as planktonic culture using Nutrient Broth No. 2 (NB2) and sessile culture (NB2+agar) and resuspended in MiliQ water.
Cell surface charge determined using zeta potential (ZP) measurement. Cells were grown as planktonic culture using Nutrient Broth No. 2 (NB2) and sessile culture (NB2+agar) and resuspended in 150 mM phosphate-buffered saline (PBS) (pH 7.4).
Bacterial attachment to abiotic surfaces determined based on the number of cells attaching to stainless steel and glass. Cells were grown as planktonic culture using Nutrient Broth No. 2 (NB2) and sessile culture (NB2+agar) and resuspended in 150 mM PBS (pH 7.4). Counts of attached cells were determined using epifluorescence microscopy after 20 min attachment at 25°C.
Planktonic or sessile mode of growth affects the cell surface properties and attachment to abiotic surfaces of both C. jejuni and C. coli
For all the strains tested, cells had a significantly (p<0.05) higher water contact angle when grown as planktonic cultures than as sessile cultures, indicating that planktonic cells are more hydrophobic than their sessile counterparts using this measure. With the exception of the ΔkpsE mutants of the strains C719a and C977a, planktonic cells of all the other strains also showed significantly (p<0.05) stronger AAG activity than sessile cells after 4-h incubation. However, planktonic cells had variable surface negative charge and attachment counts to stainless steel and glass as compared to sessile cells depending on the strain (Table 3).
Discussion
The role of surface polysaccharides in affecting Campylobacter cell surface hydrophobicity, surface charge, AAG, and mediating attachment to abiotic surfaces was investigated in the present study. Campylobacter cells possess four different surface polysaccharides, namely, CPS, LOS, and O-linked and N-linked glycans (Guerry and Szymanski, 2008). To determine the contribution of CPS, the kpsE gene was inactivated. The kpsE gene is involved in the transport of the CPS polymers to the bacterial cell surface and consequently mutation of the kpsE gene results in nonencapsulated mutants (Bachtiar et al., 2007). Four ΔkspE mutants have been successfully generated in three C. jejuni strains and one C. coli strain.
Our study showed that CPS might have only a minor contribution to cell surface properties, AAG, and attachment to abiotic surfaces of the three C. jejuni strains. However, since the wild-type of two C. jejuni strains do not express CPS, it is not possible to observe the effect of CPS on the functional assays related to cell surface hydrophobicity, AAG, or attachment to abiotic surfaces of C. jejuni. This claim, therefore, needs to be validated using other strains that are able to express CPS or by changing culture conditions stimulating CPS expression by our strains. The role of CPS in affecting cell surface properties and AAG may vary depending on individual strains, since our results did not agree with that of previous studies, which reported that loss of CPS increases the surface hydrophobicity of C. jejuni 11168 (Jeon et al., 2009) but decreases AAG activity of the strain C. jejuni 81–176 (Guerry et al., 2006). The differences in surface properties between wild-type strains and isogenic mutants may be attributable to procedural differences between studies. Future work on additional strains, such as C. jejuni 81–176 and C. jejuni 11168, and using the assays as described in the previous studies may help rule out this possibility. CPS may also have little role in cell surface hydrophobicity and attachment to abiotic surfaces of the C. coli strain, but it is required to maintain the surface charge on the cell surface and is strongly involved in AAG of the strain.
Our study also showed that LOS core has little role in affecting cell surface properties, AAG, and attachment to abiotic surfaces of C. jejuni. However, LOS, unlike CPS, may not be required to maintain the surface charge and AAG of the C. coli strain but may affect cell surface hydrophobicity and attachment to abiotic surfaces of this strain. Increased cell surface hydrophobicity is known to enhance bacterial attachment to surfaces by increasing hydrophobic interactions (Bos et al., 1999; Nguyen et al., 2011). However, this is not the case in the present study since truncation of LOS core oligosaccharides rendered the C. coli strain more hydrophobic but reduced its attachment number to these surfaces. Our results support the claim that explaining bacterial attachment to inert surfaces based on nonspecific interactions such as hydrophobic and electrostatic interactions is not always successful (Parkar et al., 2001; Roosjen et al., 2006). The reduced ability of the more hydrophobic waaF mutant of the C. coli strain to attach to abiotic surfaces may be due to a reduction of specific interactions between LOS and abiotic surfaces upon truncation of LOS.
In addition to the abovementioned findings, our study also demonstrated that planktonic or sessile mode of growth may affect cell surface properties and their subsequent ability to attach to abiotic surfaces. While it is not clear which mode enhanced their surface charge and attachment to abiotic surfaces since this varied between strains, planktonic cultures displayed increased cell surface hydrophobicity and AAG activity as compared to sessile cultures. It has been shown that relative to planktonic cells, sessile cells also exhibited a repression of genes involved in motility and flagellar expression (Sampathkumar et al., 2006). Since Campylobacter hydrophobicity and AAG are strongly associated with flagellar expression (Misawa and Blaser, 2000), it is possible that sessile cells of our Campylobacter strains demonstrate a repression in flagellar expression, which subsequently renders them less hydrophobic and prone to agglutination than planktonic cells.
In conclusion, this work demonstrated that CPS and LOS have an impact on the cell surface properties, AAG, and attachment to abiotic surfaces for C. coli but not for C. jejuni. Since only a limited number of strains were studied in the present study, this claim requires further investigation using a larger number of strains of both C. jejuni and C. coli. It has been shown that hydrophobicity and AAG of C. jejuni 11168 and 81–176 are strongly associated with flagellar expression and, more specifically, O-linked flagellin glycosylation (Misawa and Blaser, 2000; Guerry et al., 2006). Further investigations on the effects of flagellin glycosylation on hydrophobicity and AAG of the C. jejuni strains in the present study are also required. This could be achieved by inactivating the pse genes, which are required for biosynthesis and/or transfer of pseudaminic acid (PseAc) and PseAm (an acetamidino form of PseAc) to flagellin, and comparing these mutants to wild-type strains with normally glycosylated flagella.
Disclosure Statement
No competing financial interests exist.