
Abstract
A survey of cold-blooded vertebrates and associated surface waters in a produce-growing region on the Central California Coast was done between May and September 2011 to determine the diversity of Salmonella. Samples from 460 amphibians and reptiles and 119 water samples were collected and cultured for Salmonella. Animals sampled were frogs (n=331), lizards (n=59), newts (n=5), salamanders (n=6), snakes (n=39), and toads (n=20). Salmonella was isolated from 37 individual animals, including frogs, lizards, snakes, and toads. Snakes were the most likely to contain Salmonella, with 59% testing positive followed by 15.3% of lizards, 5% of toads, and 1.2% of frogs. Fifteen water samples (12.6%) were positive. Twenty-two different serovars were identified, and the majority of isolates were S. enterica subsp. IIIb, with subsp. I, II, and IIIa also found. The serovar isolated most frequently was S. enterica subsp. IIIb 16:z10:e,n,x,z15, from snakes and frogs in five different locations. S. enterica subsp. I serovar Typhimurium and the monophasic I 6,8:d:- were isolated from water, and subspecies I Duisburg and its variants were found in animals and water. Some samples contained more than one type of Salmonella. Analysis of pulsed-field gel electrophoresis pulsotypes indicated that some strains persisted in animals and water collected from the same location. Sixty-six isolates displayed antibiotic resistance, with 27 isolates resistant to more than one antibiotic, including a subspecies IIIb isolate from snake having resistance to five different antibiotics. Twenty-three isolates were resistant to more than one class of antibiotic, and six isolates were resistant to three classes. While these subspecies of IIIa and IIIb cause fewer instances of human illness, they may serve as reservoirs of antibiotic resistance, determinants in the environment, and be sources of contamination of leafy greens associated with product recalls.
Introduction
A
Wildlife is a known carrier of foodborne pathogens, with wild pigs shown to harbor Escherichia coli O157:H7 and Salmonella (Jay et al., 2007; Wacheck et al., 2010; Gorski et al., 2011). An E. coli O157:H7 strain isolated from a feral pig in Central California had the same multiple-locus variable number tandem repeat analysis (MLVA) pattern as a strain that caused an outbreak due to contaminated spinach grown in the region (Jay et al., 2007). A recent survey in that same region of soil, cattle, water, and wildlife for Salmonella showed an incidence of 2.3%. Of 240 wildlife samples tested in that survey, 4.2% were positive, including samples from birds, rodents, coyotes, elk, and deer; no amphibians or reptiles were sampled (Gorski et al., 2011). Produce grown in this region has been associated with multiple recalls due to Salmonella contamination (FDA, 2009, 2010; Anonymous, 2012; FDA, 2012a). The potential for wild amphibians and reptiles in this leafy green production region to harbor Salmonella has not been investigated. We initiated a survey in the region to assess the carriage of Salmonella by these animals and the types of Salmonella they carry in order to determine the risk of contamination of preharvest produce. In this article, we describe the characterization of Salmonella isolates from the survey including pulsed-field gel electrophoresis (PFGE) pulsotypes, serovar, and with the increased focus on multidrug resistant (MDR) Salmonella, antibiotic resistance profiles (WHO, 2005).
Materials and Methods
Sampling and enrichment for Salmonella
Animals were collected between May and September 2011 at 21 different sites in Monterey and Santa Cruz Counties of California up to 60 miles apart, and locations were assigned a seven-to-eight-character code. Hand grabs and nets were used to capture live animals from passive traps of coverboard and PVC pipe. These traps take advantage of the natural tendency of amphibians and reptiles to seek refuge, and the animals are free to enter and exit at will. Up to three samples were taken from each animal: a cloacal swab (inside the cloaca), and a ventral swab (externally from the cloaca to the neck region), and a wash. The wash was done by placing the animal in a sterile Whirl-Pak bag containing 0.5 L of phosphate buffered saline (PBS) for 10 min or until the animal defecated. Animals too small to be swabbed were pooled in groups of 10–20 in a PBS bath. After sampling, all animals were released in accordance with state wildlife agency permit and institutional animal use and care committee protocol. A paired 1-L surface water sample was collected at each sampling event. Because of the multiple sampling of individuals, 460 animals were trapped to make up 1025 samples. With the addition of 119 water samples, the sample total was 1144.
Swabs were placed into 50 mL of tryptic soy broth (TSB, Difco, Franklin Lakes, NJ). Water and PBS bath samples were filtered through a 0.45-nm membrane, and the membrane was placed into 100 mL of TSB (for turbid samples, multiple membrane units were used). The TSB cultures were grown as previously described by incubation for 2 h at 25°C, 8 h at 42°C, and holding at 4–6°C until processed for Salmonella detection (Cooley et al., 2007; Gorski et al., 2011).
Enrichment for Salmonella was done concurrently on the same TSB cultures in two different laboratories (University of California Davis and Agricultural Research Service, Albany) by two described methods (Kalchayanand et al., 2009; Gorski et al., 2011). Briefly, the TSB enrichments were plated onto modified semi-solid Rappaport Vasilliadis medium and resulting motile growth was sampled and streaked onto xylose desoxycholate agar (XLD). The TSB cultures were also subjected to immunomagnetic separation with Dynal anti-Salmonella beads (Invitrogen, Grand Island, NY) according to manufacturer's instructions; the beads were inoculated into Rappaport-Vasilliadis soya peptone broth (Oxoid, Remel, Inc., Lenexa, KS) and resulting growth was streaked onto XLD. At the Agricultural Research Service laboratory, up to five black colonies were picked from the XLD plates and confirmed as Salmonella via PCR directed against invA (Gorski et al., 2011). At the Davis laboratory, up to six black colonies from XLD were confirmed biochemically to be Salmonella with media to determine lysine utilization (lysine decarboxylase broth), citrate utilization (Simmons citrate agar), and reactions in triple sugar iron agar and urea broth. Isolates confirmed biochemically were subsequently confirmed with invA PCR.
Differentiation of Salmonella isolates
To determine whether colonies selected from a sample were siblings, they were screened with repetitive-element polymerase chain reaction (rep-PCR) with ERIC-1R and ERIC-2 primers as described (Albufera et al., 2009). The PCR reactions contained 0.3 μM of each primer, 200 μM deoxynucleotides, 2.5 U Taq polymerase (New England Biolabs, Ipswich, MA), and template was a portion of Salmonella colony. PCR conditions were as described, except the initial melt was 95°C for 6 minutes (Albufera et al., 2009). The amplicons were electrophoresed on a 20-cm 1.5% agarose gel at 120 V for 5.5 h. Rep-PCR patterns were compared, and colonies representing each pattern for a given sample were selected for PFGE analysis with XbaI as described (Ribot et al., 2006).
Images of PFGE patterns were imported into BioNumerics 6.01 (Applied Maths, Austin, TX), and bands >33 kb were assigned visually using band assignment. A standard isolate (S. Braenderup H9812) was run on each gel to allow comparisons between gels. Patterns were compared to those in our growing database of >500 Salmonella PFGE patterns. If a pattern was not a direct match to a known pattern with a previously determined serovar, then the isolate was sent to the U.S. Department of Agriculture—Animal and Plant Health Inspection Service, National Veterinary Services Laboratory (Ames, IA) for serotyping. Profile similarities were calculated using Dice binary coefficients with optimization set to 1.5%, and band position tolerances indicated in the figure legends. Cluster analysis was done using the unweighted pair group method with arithmetic averages. Uncertain bands were excluded from analysis, and a similarity matrix was calculated for the XbaI patterns.
Antibiotic resistance profiles
Isolates were assessed for antibiotic resistance by the Kirby-Bauer method using Sensi-disc susceptibility test disks (Becton-Dickinson, Franklin Lakes, NJ) deposited onto a lawn of early logarithmic phase bacterial culture in TSB swabbed onto Mueller-Hinton agar (Difco). Plates were incubated for 18 h at 37°C, the inhibition zones were measured, and resistance/susceptibility was determined. The 12 antibiotics represented seven classes, and the disc load and the threshold diameter for the zone of inhibition for resistance were as follows: amikacin (30 μg, ≤14 mm), amoxicillin/clavulanic acid (20 μg/10 μg, ≤13 mm), ampicillin (10 μg, ≤13 mm), ceftriaxone (30 μg, ≤13 mm), cephalothin (30 μg, ≤14 mm), chloramphenicol (30 μg, ≤12 mm), ciprofloxacin (5 μg, ≤15 mm), gentamicin (10 μg, ≤12 mm), kanamycin (30 μg, ≤13 mm), streptomycin (10 μg, ≤11 mm), tetracycline (30 μg, ≤14 mm), and trimethoprim/sulfamethoxazole (1.25 μg/23.75 g, ≤10 mm). E. coli ATCC strain 25922 was a control for the antibiotic resistance assay.
Results
Even though the same enrichment protocols were followed in both labs, of 1144 samples 71 were positive in one lab, 74 in the other, and 52 in both for a total of 90 Salmonella-positive samples. Because multiple samples were taken from the same individual animals, 579 individuals (including water samples) were sampled, resulting in 52 (9.0%) positive individuals (Table 1). One lab had 42 positive individuals, the other had 44, with 33 positive individuals in common between labs. Each sample from a positive individual was not necessarily positive. For the purpose of this study, a sample resulting from pooled, small animals was treated as one individual. Thirteen individuals yielded more than one type of Salmonella serovar, so the 90 positive samples resulted in 106 isolates and 22 different serovar designations.
Water sampled from ponds, catch basins, ditches, reservoirs, greenhouses, wetlands, and rivers where individual animals were found.
Some serotypes found in more than one type of sample.
Snakes had the highest percentage of Salmonella-positive individuals followed by lizards. Only 6.7% of the sampled individuals were snakes, but they represented 59.0% of the positive individuals. Lizards represented 10.2% of the total individuals, but 15.3% were positive. Frogs comprised over 50% of the samples, but only 1.2% positive individuals. Water samples, which were collected from ponds, ditches, reservoirs, and wetlands where animals were sampled, had the third highest percentage of positives at 12.6%. Ponds and water from catch basins (also called sediment basins) accounted for the majority of the positive water samples.
Subspecies and serovar analysis
The identities of the Salmonella subspecies and their serovars are shown in Table 2. In all cases, preliminary assignment of serovar based on PFGE patterns was confirmed by conventional serotyping. Most of the isolates were S. enterica subsp. IIIb, and nine serovars of this subspecies were associated with 32 individuals including 22 snakes, three frogs, one toad, one lizard, and five water samples. Eight different subsp. I serovars were identified from 16 different individuals, with the majority coming from water samples followed by lizards and snakes. One individual carried an untypable Salmonella isolate. Three serovars were identified as unknown subspecies because of autoagglutination. Most of the positive samples yielded only one type of Salmonella, but 15 individuals (not pooled animals) harbored more than one serovar. Ten of those individuals are detailed in Table 3.
These strains are variants of the Duisburg serovar.
Isolates carrying Duisburg variants were excluded from this table.
These are separate individuals at the same site on the same day.
The serovar identified most often was IIIb 16:z10:e,n,x,z15 from frog and snakes, from five different locations collected May–September. Figure 1A shows a dendrogram of the XbaI PFGE patterns of IIIb 16:z10:e,n,x,z15 strains isolated from each individual. All but one shared at least 90% similarity. There are several instances of the same pulsotype being isolated from different individuals on or near the same date from the same location (e.g., MJ643-RM15868, MJ669-MJ670, MJ658-MJ661, and RM15054-RM15056). The PFGE patterns for IIIb 50:r:z, the second most common subspecies IIIb serovar, are in Figure 1B. These isolates came from water, snakes, and toad from June to September from the same location, indicating persistence of this pulsotype over time.
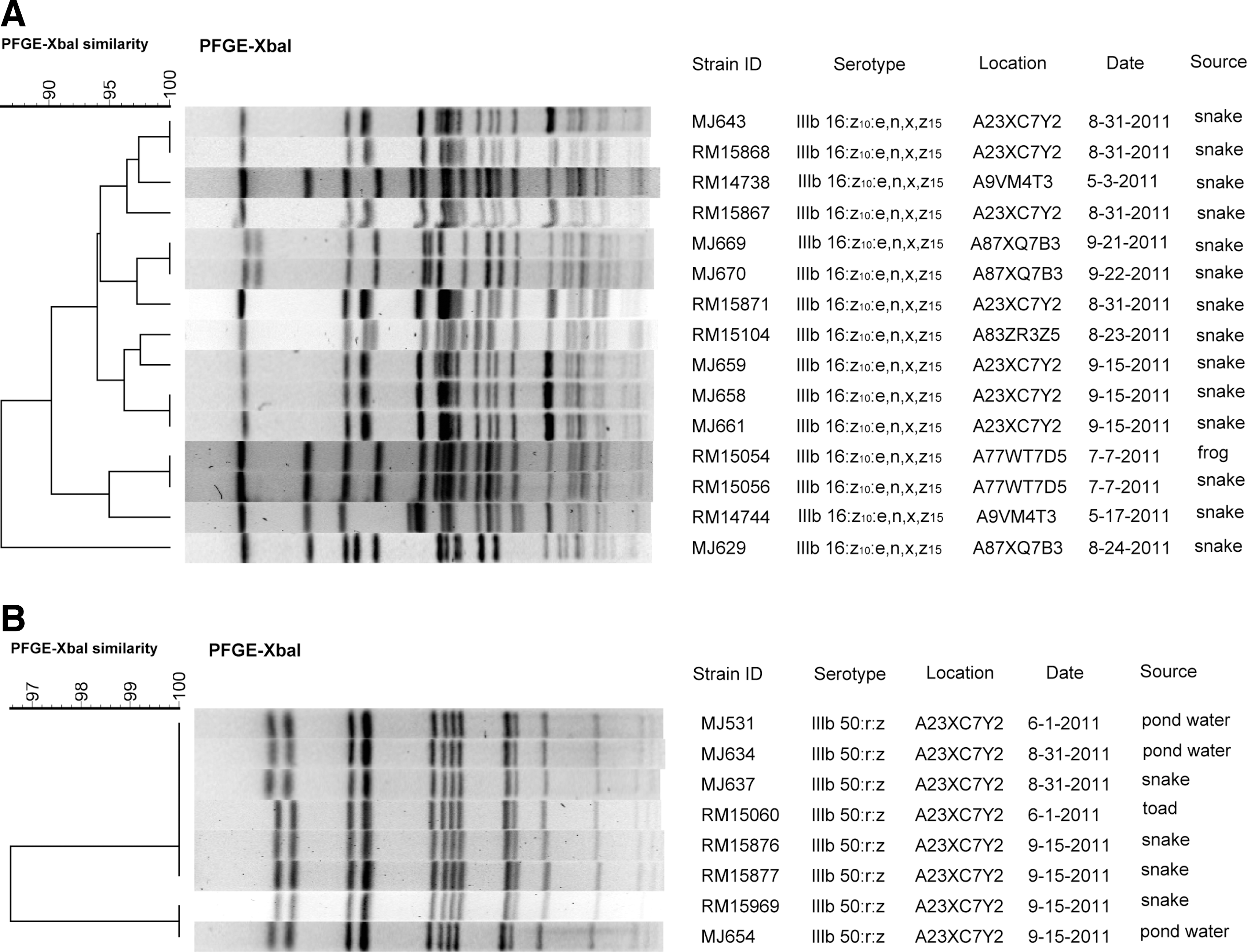
Dendrograms comparing XbaI PFGE pulsotypes of isolates of
In addition to serovar IIIb 16:z10:e,n,x,z15, other serovars isolated from more than one location were IIIa 41:z4,z23:-, and subspecies I serovars Typhimurium and the monophasic 6,8:d:- (Fig. 2). Three isolates of IIIa 41:z4,z23:- were isolated from three different locations. The isolates from sediment basin water and lizard isolated 4 months apart had >95% similar pulsotypes (Fig. 2A). Four Typhimurium isolates were from water sources from May to September at three different locations (Fig. 2B). Strains RM16181 (September) and RM14750 (May) from different locations have a >95% similarity in their pulsotypes; however, RM14750 has a Typhimurium variant serovar missing O-antigen 5. It is possible these are the same strain that underwent a phase variation. Figure 2C shows the pulsotypes for the 3 I 6,8:d:- strains that were isolated from two locations in May–September, and their pulsotypes are 85% similar. The rest of the serovars were associated with single locations, and some of these serovars were associated with multiple individuals. PFGE patterns from two of these serovars are shown in Figure 2. Isolates of IIIb 38:l,v:z53 (Fig. 2D) were found in three different snakes on the same day, and these isolates shared nearly 90% similarity with two individuals carrying the same pulsotype. Isolates of IIIb 60:r:e,n,x,z15 (Fig. 2E) came from frog, lizard, and snake, and the isolates shared 90% similarity with the frog and lizard isolates having identical pulsotypes. Not all of the isolates of serovars IIIb 59:k:z35 (two snakes) and IIIb 38:k:z35 (one frog and one snake) resolved with PFGE, so those comparisons are not shown.
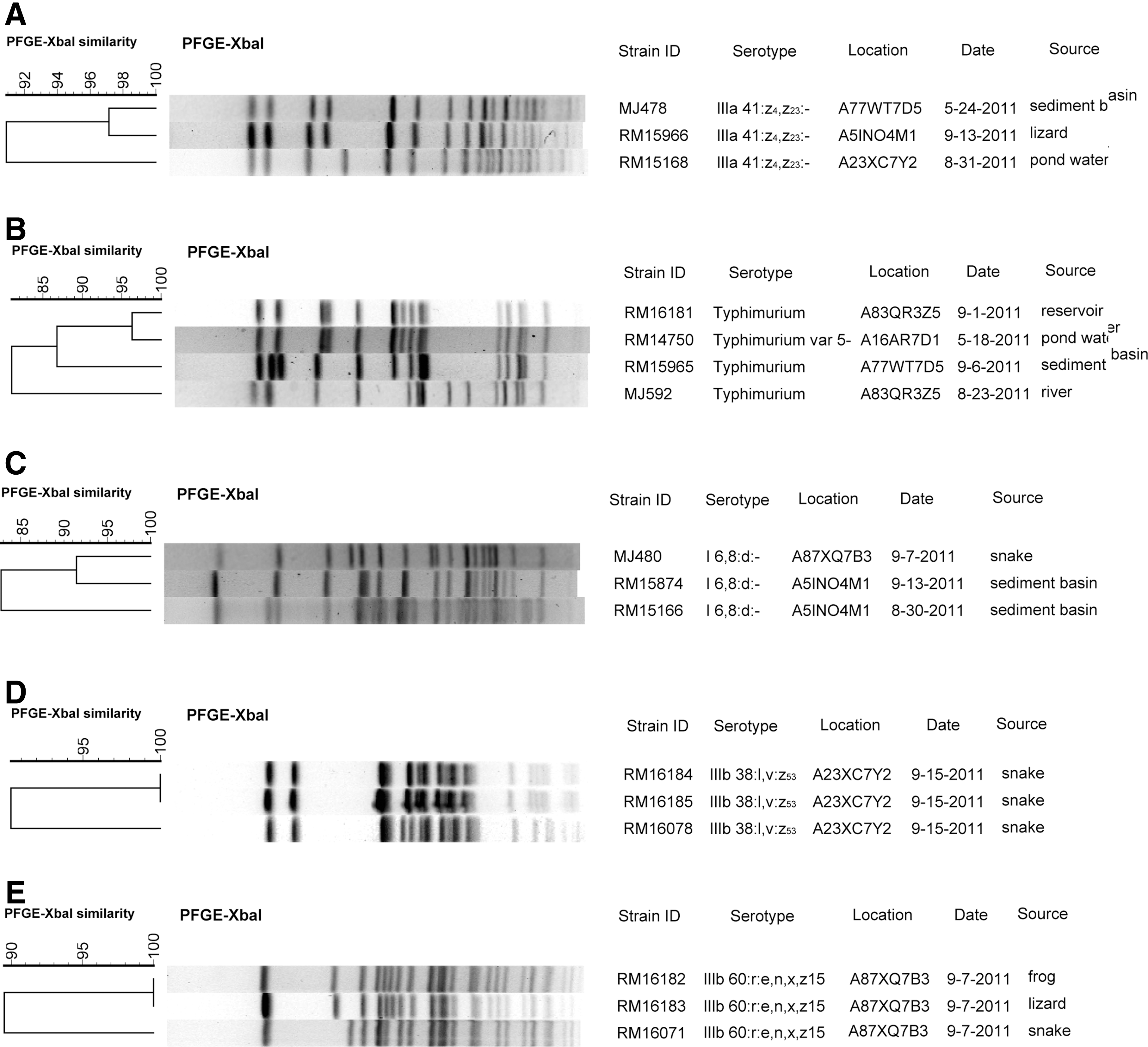
Dendrograms comparing XbaI pulsed-field gel electrophoresis (PFGE), pulsotypes of isolates of
Subspecies IIIa and IIIb isolates predominantly came from animals, while subspecies I was mostly isolated from water. The serovar isolated most frequently among subspecies I was Duisburg, for which the antigenic designation is I
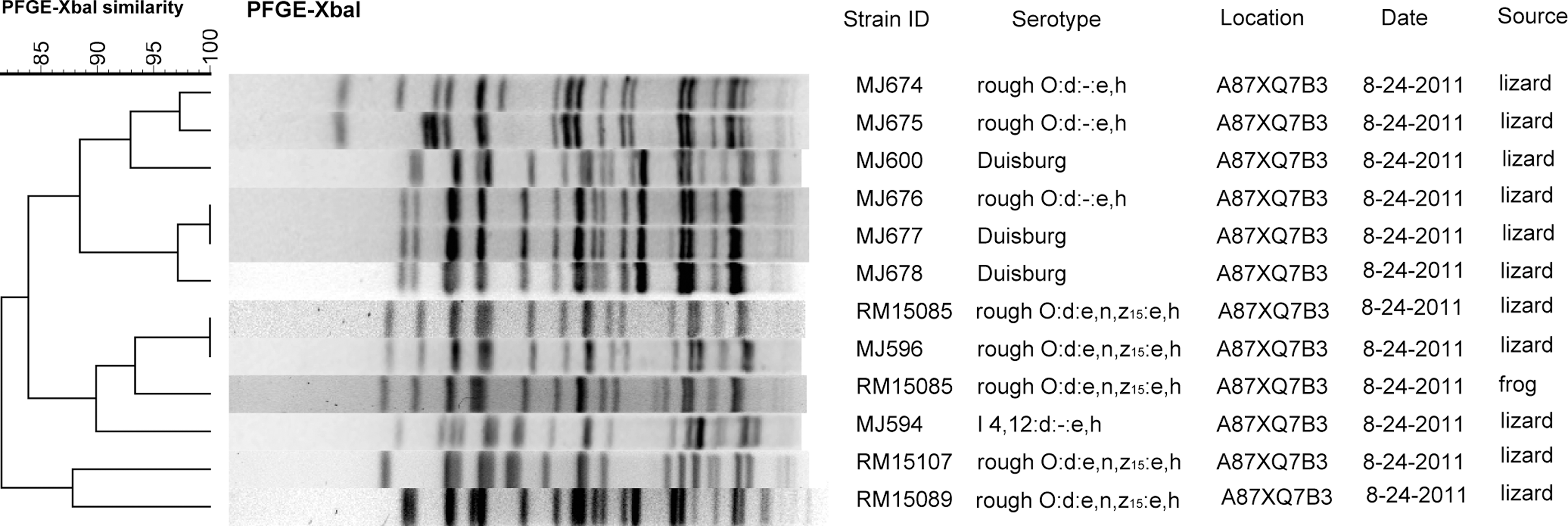
Dendrograms comparing XbaI pulsed-field gel electrophoresis (PFGE) pulsotypes of isolates of Salmonella enterica subsp. enterica strains with antigenic designations Duisburg, I 4,12:d:-:e,h; rough O:d:e,n,z15:e,h; and rough O:d:-:e,h with band position tolerance set at 4%. Columns are labeled with strain, serovar, location code, date of sample collection (month–day–year), and source. Each isolate was from a separate individual.
The remaining rough serovar roughO:-:z has a similar designation to III 50:r:z; however, the PFGE patterns indicated they were not similar (data not shown).
Antibiotic resistance profiles
All 106 isolates were tested for antibiotic resistance, and 40 were sensitive to all 12 of the antibiotics. The rest were resistant to at least one antibiotic, and 27 were resistant to more than one (Table 4). Resistant isolates came from 40 individuals (two frog, five lizard, 21 snake, one toad, and 11 water). Twenty-three isolates were resistant to more than one antibiotic class. Six isolates were resistant to three or more classes, making them MDR as defined by the National Antimicrobial Resistance Monitoring System (FDA, 2012b), including isolates of IIIb 59:k:z35 (snake), IIIb 16:z10:e,n,x,z15 (frog and snake), IIIb 50:r:z (toad), and Santiago (water). The most resistant isolate was a subsp. IIIb 38:l,v:z53 isolate (snake), which was resistant to amikacin, gentamicin, streptomycin, cephalothin, and ampicillin. Isolates were resistant most often to streptomycin (42), cephalothin (25), and ampicillin (10). All isolates were sensitive to sulfamethoxazole/trimethoprim and chloramphenicol.
Ak, amikacin; Amc, amoxicillin-clavulanic acid; Amp, ampicillin; Cro, ceftriaxone; Cf, cephalothin; Cl, chloramphenicol; Cp, ciprofloxacin; Gm, gentamicin; Km, kanamycin; St, streptomycin; Te, tetracycline.
Discussion
Of the >2400 Salmonella serovars, those with one-word, trivial names are S. enterica subsp. I. The more complex, exotic serovars or antigenic designations are associated with the other subspecies (Brenner et al., 2000; Grimont and Weill, 2007). The serovars of Salmonella found from cold-blooded vertebrates in previous studies vary by location, but complex serovars are identified frequently. The majority of isolates from a study of captive reptiles in Denmark were subspecies IIIa and IIIb (Pedersen et al., 2009). The distribution of Salmonella identified from Tokay geckos imported from Indonesia were subspecies I, IIIa, and IV (Smith et al., 2012). A recent report of Salmonella prevalence in toads in Grenada found subspecies I and IV (Drake et al., 2012).
This is the first study of the serovar distribution among wild, cold-blooded vertebrates in the Central Coast region of California. A 2009 survey of soil, water, wildlife, plants, and cattle in the same region identified mostly subspecies I and 2 isolates of subspecies IV (Gorski et al., 2011). Similar to other reptile studies but in contrast to the 2009 environmental study, the serovars identified here mostly were from subspecies IIIa and IIIb. Two serovars found in both studies were Typhimurium and the monophasic subspecies I strain 6,8:d:-. In this study, Typhimurium and the monophasic I 6,8:d:- were found only in water samples, similar to the previous study (Gorski et al., 2011). Serovar Duisburg has been found in surface and drinking waters in Europe, but we found it here associated only with lizards (Rolland and Block, 1980; Müller and Mossel, 1982).
One difference between the current and previous studies of the region is the high amount of antibiotic resistance among isolates in the current study. Only one isolate from the 2009 study was antibiotic resistant. Of particular concern is the subspecies IIIb isolates having resistance to more than two different classes of antibiotics. Other surveys of Salmonella in California have identified MDR strains of serovars Agona, Anatum, Newport, and Reading from cattle (Berge et al., 2004; Brichta-Harhay et al., 2011).
Human illness from subtype IIIa and IIIb strains is less common; however, invasive disease and death among pediatric and immunocompromised patients are associated with subspecies IIIa due to pet reptile exposure (Mahajan et al., 2003; Kolker et al., 2012). Katribe et al. (2009) reported that subspecies IIIa strains had poor colonization of mouse intestines, and while subspecies IIIb strains did colonize, they did not become systemic. However, certain IIIa and IIIb serovars have been associated with increased virulence for humans (Weiss et al., 1986). An analysis of data from 1967 to 1976 found human infections from many subspecies III serovars, including some identified in this study [IIIb 16:z10:e,n,x,z15, IIIb 38:(k),z35, IIIb 38:l,v:z53, IIIb 50:l,v:z35, and IIIb 60:r:e,n,x,z15]. Also, serovars IIIb 50:l,v:z35 and IIIb 16:z10:e,n,x,z15 were isolated from food or food products (Weiss et al., 1986). Recently, Abbott et al. (2012) reported an increase in infections, including extraintestinal infections, caused by subspecies II, III, and IV strains in California in a retrospective analysis of the years 1985–2009. Produce contaminated with these serovars may cause human illness. More research is needed to determine whether these serovars are likely to interact with produce.
Our results indicate that subtype IIIa and IIIb strains may be a reservoir of antibiotic resistance. Studies in Denmark, Germany, and of imported Indonesian geckos identified increased antibiotic resistance among the less common subspecies I serovars (Frech et al., 2003; Bagger-Skjøt et al., 2007; Smith et al., 2012). It is possible the antibiotic resistance determinants responsible for the widespread antibiotic resistance we report here could be transferred to other bacteria, including non-salmonellae, via horizontal gene transfer. Samples yielding multiple types of Salmonella indicate that several types can coexist in the same environment and transfer resistance genes potentially to other strains (Allen et al., 2010). Other studies have reported reptiles carrying more than one type of Salmonella (Willis et al., 2002).
A variety of salmonellae were isolated in this study, but it is possible that others were present and undetectable by the methods used. Enrichment media can bias the types of Salmonella isolated (Singer et al., 2009; Gorski, 2012; Taskila et al., 2012). One advantage to testing in two laboratories using two different Salmonella enrichment techniques was to minimize culture bias and increase chances of isolating different types of Salmonella. More samples were positive when the results from the labs were compiled than from either lab by itself.
Another finding of note was the isolation of Salmonella from sediment basins and grassed ditches. These water sediment samples were contaminated frequently with subspecies I strains, which are more common in human illness than the other subspecies. Thus, the incidence of potential human pathogens in cold-blooded vertebrates associated with the region and recent recalls associated with Salmonella-contaminated leafy greens from this region emphasize the need for growers to be made aware of this potential source of preharvest contamination.
Conclusions
Salmonella enterica subsp. I, II, IIIa, and IIIb were isolated from amphibians and reptiles in a produce-growing region of the Central California coast. Many of the isolates were of subtypes IIIa and IIIb, which are seen less commonly in human infections than subspecies I. The presence of antibiotic resistance in a majority of the isolates indicates that they could be a source of antibiotic resistance determinants in the environment.
Footnotes
Acknowledgments
We are grateful to Dawn Reis, Jessica Wheeler, Jordi Montfort, Alexis Fisher, Stephanie Huang, Diana Carychao, and Kimberly Nguyen for technical assistance. We also extend our thanks to the growers and property owners who kindly allowed access to their farms. Funding for this project was provided by the Center for Produce Safety through the California Department of Food and Agriculture, Specialty Crop Block Grant #SCB10056, the U. S. Food and Drug Administration (Project No. U01-003-572), and U.S. Department of Agriculture, Agricultural Research Service CRIS project 5325-42000-046-00D.
Disclosure Statement
No competing financial interests exist.