
Abstract
Vibrio parahaemolyticus is a major foodborne pathogen in China and other countries. In this study, a total of 578 clinical V. parahaemolyticus strains and 51 foodborne strains were isolated during the period from 2009 to 2011 in the eastern coastal city of Shanghai, China. Their serotypes, virulence genes, pandemic traits, and genotyping were investigated. A total of nine O groups and 20 K types were identified by serological analysis of all isolates. Six different O groups and 14 different K types were detected among the 578 clinical strains. Eight different O groups and five K types were detected among the 51 foodborne strains. The O3:K6 serotype was the dominant serotype. A total of 200 representative clinical strains and 51 foodborne isolates were analyzed for virulence genes, pandemic traits, and genotyping. Of the clinical strains, 92.5% had the virulence genes tdh and/or trh. Four foodborne isolates had virulence genes; one trh-positive strain was O3:K6 and three tdh-positive strains were either O4:KUT or O3:KUT. Molecular typing by pulsed-field gel electrophoresis also showed divergence among the nonpandemic strains, although the pandemic strains formed a cluster. These results suggest high serodiversity and genetic diversity of V. parahaemolyticus. Pathogenic isolates were present in food, thus representing a public health risk and warranting epidemiological and ecological monitoring to ensure safety.
Introduction
Previously, cases of V. parahaemolyticus infections have typically been sporadic and attributed to multiple serotypes, with at least 13 O serogroups and 71 K serotypes detected (Ishibashi et al., 2000). However, outbreaks of V. parahaemolyticus infections that occurred after 1996 were initially linked to a predominant serotype O3:K6. This “new” O3:K6 first appeared in 1996 in Kolkata, India (Okuda et al., 1997) and rapidly spread worldwide. An increasing incidence of gastroenteritis caused by the “new” O3:K6 has been reported in many regions, including Africa (Ansaruzzaman et al., 2005), Europe (Martinez-Urtaza et al., 2004), Latin America (González-Escalona et al., 2005), Mexico (Velazquez-Roman et al., 2012), and Chile (Harth et al., 2009). Serotype O3:K6 was then identified as a dominant pandemic clone isolated from clinical cases of diarrhea reported globally (Okuda et al., 1997; Smolikova et al., 2001). To date, at least 21 other serovariants have been identified in addition to the original O3:K6 serotype (Nair et al., 2007).
It has been shown that pandemic V. parahaemolyticus strains can be identified by group-specific polymerase chain reaction (GS-PCR) based on sequence variation in the toxRS gene (toxRS/new) that is unique to the pandemic O3:K6 clone (Okura et al., 2003). Therefore, strains that are positive for both tdh and GS-PCR can be reliably identified as pandemic strains (Matsumoto et al., 2000).
V. parahaemolyticus was originally identified as the causative agent of a gastroenteritis outbreak in Japan in 1950 (Fujino et al., 1951). As one of the most important foodborne pathogens in China, V. parahaemolyticus was first identified in Shanghai in 1958. Pathogenic V. parahaemolyticus accounts for 63.7% of cases of food poisoning in Shanghai each year (Liu, 2003). However, less information is available with regard to the properties of V. parahaemolyticus isolated from patients and food in the coastal area of Shanghai, China. Therefore, the main purpose of this study was to compare V. parahaemolyticus isolates in terms of serotype, virulence genes, dissemination of pandemic clones and genotypic traits, in order to provide useful information that can be used by government agencies to perform risk analysis and to better control the dissemination of this pathogen.
Materials and Methods
Bacterial strains
A total of 629 V. parahaemolyticus strains were used in this study, including 578 clinical strains and 51 foodborne isolates obtained from 2009 to 2011 in Shanghai. Overall, 578 clinical strains were isolated from outpatients with sporadic diarrhea from the following active surveillance hospitals: Putuo (78), Changning (23), Yangpu (48), Zhabei (17), Nanhui (30), Hongkou (100), Luwan (75), Xuhui (14), Baoshan (162), Fengxian (10), and Qingpu (21). Fifty-one food strains were isolated from aquatic products obtained from retail markets, supermarkets, restaurants, and hotels. Of these, 24 strains were from fish, 16 strains from shrimp, eight strains from shellfish, and three strains from crab. With regard to the geographic sources of the 51 foodborne isolates, five strains were isolated from Fengxian, eight strains from Nanhui, four strains from Baoshan, four strains from Pudong, 12 strains from Yangpu, seven strains from Changning, three strains from Putuo, six strains from Jinshan, and two strains from Luwan.
All strains were isolated and identified as described in GBT 4789.7-2008. Briefly, the strains were inoculated onto thiosulfate–citrate–bile salt–sucrose agar with prior enrichment culture in alkaline peptone water, and were incubated (18–24 h) at 37°C. The strains were identified as V. parahaemolyticus by standard biochemical tests. Two ATCC strains—ATCC 33847 (tdh +) and ATCC17802 (trh +)—were used as positive controls in this study.
Serotyping
Serotyping of V. parahaemolyticus isolates was conducted using a commercially available V. parahaemolyticus antiserum test kit (Denka Seiken, Tokyo, Japan) in accordance with the instructions provided by the manufacturer. Briefly, strains were cultured overnight at 37°C on Luria-Bertani agar containing 3% NaCl. A pool of colonies was then collected and resuspended in 1 mL normal saline. An aliquot (0.5 mL) was sterilized by autoclave (121°C for 1 h) and lysates were then used for serotyping based on the O antigen. The strains that did not agglutinate with O antisera were sterilized by autoclave for 2 h, and the remaining unsterilized cell suspension was used for serotyping based on the K antigen.
PCR assays
The toxR gene was used to identify the V. parahaemolyticus strains. The toxR gene sequences have been cloned from V. cholerae, V. fischeri, and at least two other Vibrio species (DiRita, 1992; Reich et al., 1994). The specific sequence of the toxR gene was used to detect V. parahaemolyticus. The two virulence genes tdh and trh were used to detect pathogenic V. parahaemolyticus. The PCR assays for the species-specific gene, toxR, and two virulence genes (tdh, trh) were performed using V. parahaemolyticus genomic DNA as a template, following previously described methods (Bej et al., 1999). GS-PCR, a method to specifically detect the toxRS sequence (toxRS/new) of the pandemic clone in V. parahaemolyticus, was performed using the specific primers that were reported by Matsumoto (Matsumoto et al., 2000). All primers used are described in Table 1.
Forward primer.
Reverse primer.
GS-PCR, group-specific polymerase chain reaction.
PFGE
It has been reported that the PFGE analysis showed a high degree of clonality among V. parahaemolyticus isolates following digestion of DNA with NotI and SfiI (Martinez-Urtaza et al., 2004). Kam (2008) also recommended the use of the SfiI as the primary enzyme. Therefore, V. parahaemolyticus isolates were digested using SfiI. Pulsed-field gel electrophoresis (PFGE) of DNA obtained from V. parahaemolyticus was performed using a standard protocol with a little modification (PulseNet, 2009). XbaI-digested Salmonella Braenderup H9812 DNA was used as molecular size markers. Electrophoresis was performed using the CHEF-DRII apparatus (Bio-Rad Laboratories, Hercules, CA). Images were captured on the Gel Doc 2000 system (Bio-Rad Laboratories) and converted to TIFF files for computer analysis.
Results
Serotyping analysis
Nine different O groups and 20 different K types were identified among 629 clinical strains and foodborne isolates (Table 2). Among the 578 clinical strains, six different O groups and 14 different K types were detected. Fifteen strains were not recognized by specific O antisera, and 89 strains were not recognized by specific K antisera. These strains may be new serotypes. Of the 578 clinical strains, the most frequent O group was O3 (384), followed by O4 (117) and O1 (56), O5 (2), O2 (2), and O8 (2). A total of 14 K types were detected representing 25 different serotypes. The dominant serotype was O3:K6, accounting for 360 strains (62.2%), followed by O4:K8 (63), O4:K68 (25) and O4:K9 (13). Other O:K serotypes included O1:K56 (9), O4:K4 (3), O1:K36 (4), and O4:K55 (2). Furthermore, O3:K20, O3:K9, O4:K13, O4:K12, O2:K28, O1:K8, O3:K8, O4:K37, O4:K56, and O3:K5 serotypes were each represented by one strain. Of the 578 strains belonging to serogroup O, for which the K types were not typeable (i.e., did not react with any of the available K antisera), the following serotypes were identified: O1:KUT (42), O3:KUT (21), O4:KUT (5), O8:KUT (2), O5:KUT (2), and O2:KUT (1). Newly emerging serotypes were detected including O1:K8 (1), O3:K20 (1), O4:K37 (1), and O1:K36 (4). In contrast, a decreasing frequency of O3:K6 strains (from 65.4% to 47.1%) was observed between 2009 and 2011 (Fig. 1). There were sharp increases in O4:K68, O4:K9, and O3:KUT serotypes (from 3.21% to 9.24%, 0.64% to 5.88%, and 1.92% to 11.76%, respectively) over the same period (Fig. 1).
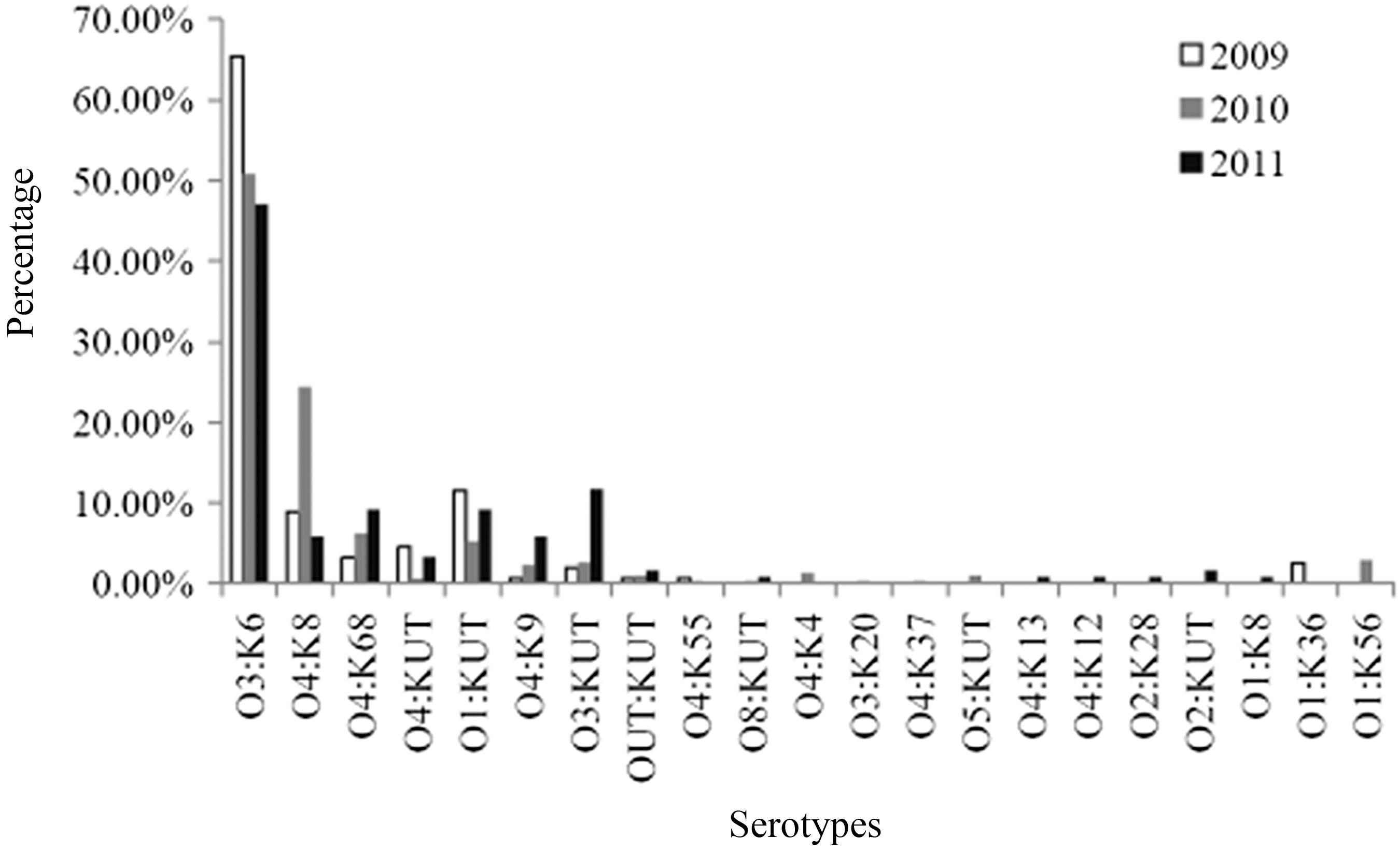
Distribution of serotypes of clinical Vibrio parahaemolyticus strains isolated in Shanghai during the period from 2009 to 2011.
Newly emerging serotypes.
The serotypes of food isolates of V. parahaemolyticus differed from those of the clinical strains (Fig. 2). Eight different O groups were detected. The predominant O groups were O3 (16), followed by O1 (10), O4 (7), and O5 (7). Other O groups included O2 (2), O10 (2), O11 (2), and O6 (1). However, most strains did not react with K antisera. Approximately 11.8% of 51 strains isolated from food were typed by K antisera as O3:K6, O4:K42, O4:K8, OUT:K2, O2:K28, and O1:K33 (1 strain each). O1:K33 was identified as a newly emerging serotype. The same O:K serotypes (O3:K6, O4:K8, O2:K28) were detected in clinical strains.
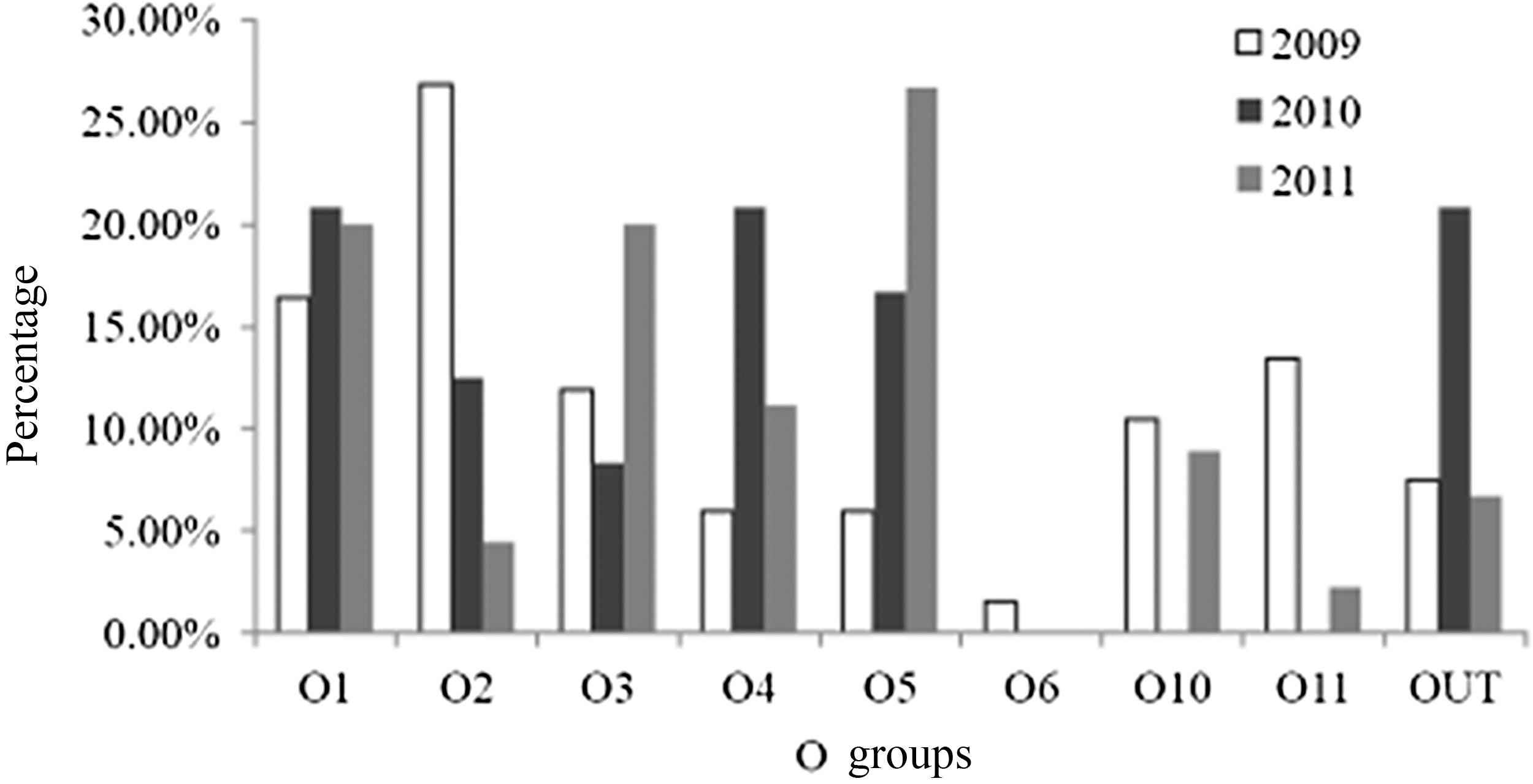
Distribution of serotypes of foodborne isolates of Vibrio parahaemolyticus in Shanghai during the period from 2009 to 2011.
Detection of virulence genes by PCR
Representative strains, including 200 clinical strains and 51 foodborne isolates, were selected for further study. Selection was made in order to represent a variety of different patterns including dissimilar serotypes and periods of isolation. The species-specific toxR gene was detected in all 251 V. parahaemolyticus strains (Table 3). Of 200 clinical strains of V. parahaemolyticus, 185 (92.5%) strains had the tdh and/or trh virulence genes, of which 172 (93%) strains had the tdh virulence gene. Of these strains, 13 had trh but not tdh, and six strains had tdh and trh. The clinical strains with tdh and/or trh belonged to 24 serotypes, most of which were O3:K6 (58/58), O4:K8 (35/38), O4:K68 (21/21), O1:KUT (15/18), followed by O3:KUT (12/15), O4:K9 (11/13), O4:KUT (10/10), O1:K36 (3/3), O4:K4 (3/3), O4:K55 (2/2), O1:K56 (2/2), O2:K28 (1/1), O1:K8 (1/1), O5:KUT (1/1), O3:K8 (1/1), O3:K9 (1/1), O8:KUT (4/4), O3:K5 (1/1), O4:K56 (1/1), O3:K20 (1/1), and OUT:KUT (1/1). In contrast, other serotypes, including O4:K12, O4:K13, OUT:K24, and O1:K33, had no virulence genes. However, 47 (92.2%) foodborne V. parahaemolyticus strains isolates did not carry virulence genes, although one strain (detected in fish) had the trh gene and was serotype O3:K6; three strains (two from fish, one from shrimp) had the tdh gene and were serotype O3:KUT and O4:KUT, respectively.
Three strains from fish, two strains from shellfish, eight strains from shrimp, two strains from crab.
One strain from fish, one strain from shrimp, one strain from shellfish.
Three strains from fish, two strains from shrimp, three strains from shellfish, one strain from crab.
Two strains from fish, one strain from shrimp.
GS-PCR, group-specific polymerase chain reaction.
Detection of pandemic marker genes by PCR
As shown in Table 2, 29% (58/200) of clinical strains were positive for tdh and GS-PCR, which is characteristic of pandemic clones. Of the 58 pandemic strains, 35 (60.3%) strains belonged to O3:K6, followed by O4:K68 (11), O1:KUT (5), O3:KUT (2), O1:K36 (3), O1:K56 (1), and O3:K8 (1). It is noteworthy that pandemic marker genes were not detected in the 51 foodborne strains.
PFGE and cluster analysis
In this study, 251 strains isolated from patients and food were typed by PFGE analysis. As shown in Figure 3 (partial data shown), different PFGE patterns were obtained for V. parahaemolyticus strains. First, these observations indicate that strains of the same serotypes and having a similar virulence gene profile produced the same or similar PFGE patterns. For example, two O4:K8 strains (SH2010047 and SH2010048, clinical strains) with no virulence genes belonged to the same cluster. Second, strains of different serotypes had the same or similar PFGE patterns. For example, O4:K8 (SH2010031) and O4:K68 (SH2010068) strains had the same PFGE patterns. Third, the PFGE patterns of V. parahaemolyticus strains from different sources were the same or very similar. For example, one O3:KUT strain (SH201181) isolated from shrimp had the same PFGE patterns as one O3:KUT clinical strain (SH201183). The PFGE patterns of one food pathogenic isolate (SH20112916) and three clinical pathogenic isolates are very similar (Fig. 3). Not unexpectedly, the pandemic strains clustered together (Fig. 4). The cluster constituted mainly pandemic O3:K6 serotype strains and also included O4:K68, O1:KUT, O3:KUT, O1:K36, O1:K56, and O3:K8 strains.
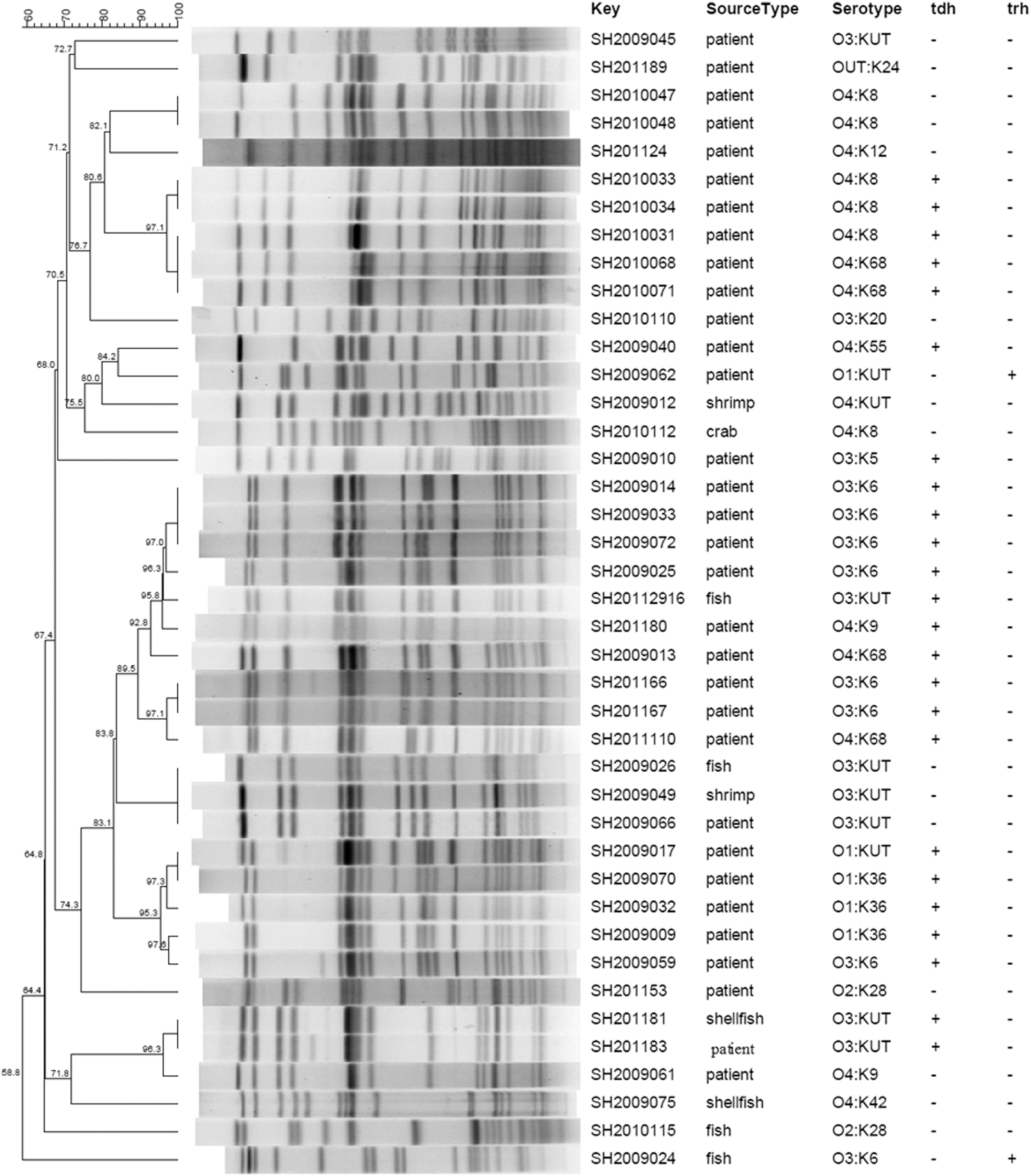
Pulsed-field gel electrophoresis patterns of SfiI-digested genomic DNA of selected clinical and foodborne Vibrio parahaemolyticus isolates obtained in Shanghai. Strain identification number, seromarkers, source of isolation, and virulence genes (tdh, trh) are indicated.
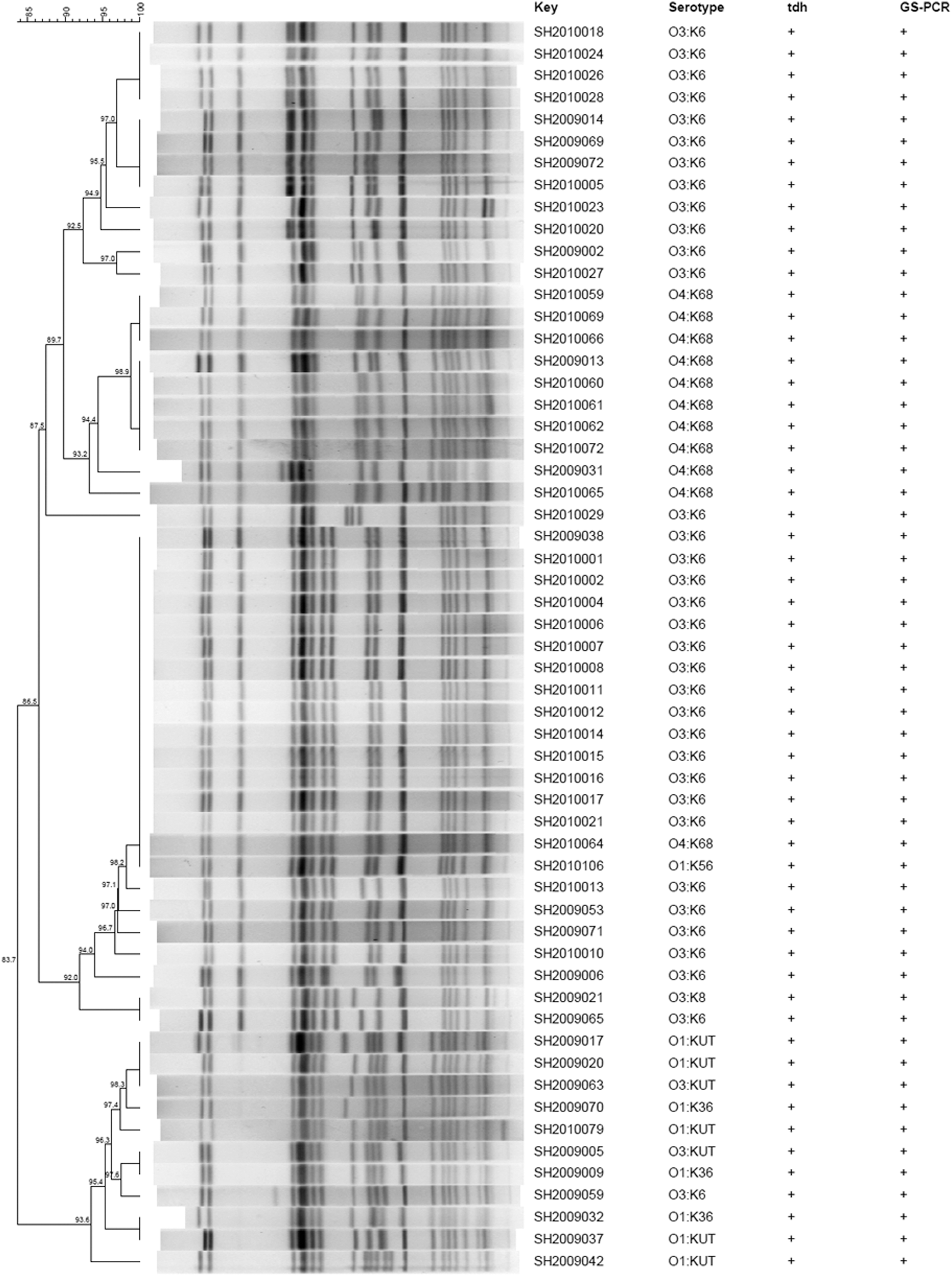
Dendrogram constructed from pulsed-field gel electrophoresis images of SfiI-digested genomic DNA of pandemic strains obtained from clinical isolates. Strain identification number, seromarkers, and pandemic strain genes including tdh and group-specific polymerase chain reaction (GS-PCR) are indicated.
Discussion
V. parahaemolyticus has emerged as a worldwide pathogen that causes gastroenteritis related to the consumption of seafood (Daniels et al., 2000; Wong et al., 2000). In recent years, new cases have arisen in China (Liu et al., 2006). This study includes data collected over the 3 years from 2009 to 2011 in Shanghai. In both clinical and foodborne isolates of V. parahaemolyticus, we identified the serotypes, virulence genes, and pandemic strains. Molecular typing of these strains was conducted by PFGE.
Serodiversity was detected among the 31 serotypes of clinical and foodborne isolates of V. parahaemolyticus. The clinical strains belonged to 25 serotypes, a diversity of strains far greater than that observed previously in other countries. In Mexico, only 14 serotypes of clinical strains were detected from 2004 and 2010 (Velazquez-Roman et al., 2012). However, in this study, a large number of foodborne isolates were untypable with the K antigen, and a few foodborne and clinical isolates were untypable with the O and K antigens, which has been reported in previous studies (Mahmud et al., 2007). Furthermore, four newly emerging serotypes that were not listed by the Japanese committee were identified based on the serological typing of V. parahaemolyticus. These were O1:K8 (clinical isolate), O3:K20 (clinical isolate), and O1:K36 (clinical isolates) and O1:K33 (foodborne isolate). The O1:K33 serotype had previously been reported in India in 2002 (Pal et al., 2010), while the other three serotypes have been reported in China (Chao et al., 2009).
The results of this Shanghai-based study showed that O3:K6 strains were dominant, accounting for 62.2% of the serotypes, which is in accordance with previous studies from Beijing, Guangdong, and Mexico (Ke et al., 2011; Qu et al., 2011; Velazquez-Roman et al., 2012). However, this frequency was lower than that observed in other countries; for example, O3:K6 accounted for 80.5% of serotypes isolated in Northern Mexico between 2004 and 2010 (Velazquez-Roman et al., 2012). Furthermore, there was a decline in O3:K6 strains in Shanghai from 2009 to 2011 (Fig. 1). This decrease may be associated with increases in other serotypes, such as O4:K68, O4:K9, and O3:KUT, which have also been reported in other countries (Nair et al., 2007; Harth et al., 2009).
Previous epidemiological studies have revealed that pathogenicity correlates strongly with two well-characterized virulence genes, tdh and/or trh (Nishibuchi et al., 1995). Therefore, the presence of these two genes is routinely used to determine the pathogenicity of V. parahaemolyticus strains. As shown in Table 3, 92.5% of clinical strains carried the tdh and/or trh gene, as has been reported previously (García et al., 2009). In contrast, virulence genes are reported to be rarely present in foodborne isolates (Chao et al., 2009), which is consistent with the findings of our studies, in which only four (7.8%) foodborne strains carried the tdh or trh genes. Serotypes of the four strains with virulence genes, O3:K6, O4:KUT, and O3:KUT, have also been detected in clinical strains. It is worth noting that the tdh-positive O3:KUT strain isolated from shrimp has the same PFGE pattern as the tdh-positive O3:KUT clinical strain (Fig. 3). Therefore, it can be concluded that the V. parahaemolyticus serotype, O3:KUT, recognized as a foodborne diarrheal pathogen in Shanghai, poses a public health risk that warrants epidemiological and ecological monitoring.
The rapid emergence of non-O3:K6 serotypes carrying pandemic markers provides another example of the predisposition of V. parahaemolyticus to genetic change (Matsumoto et al., 2000; Chowdhury et al., 2004). Six pandemic serotypes were found in clinical strains in this study in addition to O3:K6. Furthermore, the newly emerging O1:K36 pandemic serotype found in this study has not been reported in countries other than China (Chao et al., 2009). We suggest that, due to changes in the serotypes of pandemic strains, continuous monitoring of these pandemics strains will be crucial in the development of strategies to control diarrheal illness in Shanghai.
Molecular typing by PFGE was performed to assess the genetic diversity of all isolates. Our data showed great divergence among V. parahaemolyticus foodborne and clinical isolates, which is in agreement with many studies from other regions (Chao et al., 2010). Furthermore, PFGE profiles of the pandemic strains showed the same or closely related clusters. Therefore, the seromarkers currently used to categorize pandemic serotype strains of V. parahaemolyticus are of limited or no use. However, the overall PFGE and cluster analysis data, coupled with results of other investigations, provide clear evidence that PFGE is a more efficient technique for the analysis of epidemiology and risk assessment of V. parahaemolyticus infections.
Conclusions
The serotypes of 629 foodborne and clinical isolates showed high diversity. Overall, 92.5% of 200 clinical strains had virulence genes and 29% clinical strains were identified as pandemic strains. The pandemic clonal serotypes included O3:K6, O4:K68, O1:KUT, O3:KUT, O1:K56, O3:K8, and the newly emerging serotype, O1:K36. Molecular typing using PFGE showed a high rate of divergence among nonpandemic strains and pandemic clones that exhibited identical PFGE patterns. Pathogenic but not pandemic strains were detected in food, and four foodborne isolates carried the virulence genes. One food pathogenic isolate is somehow related to the clinical pathogenic strains based on PFGE typing. As a result, these pathogenic isolates were found to be present in food, posing a public health risk and warranting epidemiological and ecological monitoring to ensure safety.
Footnotes
Acknowledgments
This work was supported by a grant from the Shanghai Municipal Natural Science Foundation (No. 11ZR1431000, to Hongzhi Zhang).
Disclosure Statement
No competing financial interests exist.