
Abstract
This study presents evidence that the pattern (diffuse or aggregative) of adherence of clinically relevant non-O157 Shiga toxin–producing Escherichia coli (STEC) to bovine recto-anal junction squamous epithelial cells is similar to that of E. coli O157, although the mechanisms of adherence appear to be distinct. Our results further suggest that novel adhesins, and not Intimin, are likely involved in non-O157 STEC adherence to bovine recto-anal junction squamous epithelial cells. These findings have important implications for the development of efficacious modalities for blocking adherence of non-O157 STEC to bovine gastrointestinal epithelial cells.
Introduction
S
Ruminants (cattle and sheep) are primary reservoirs for STEC and hence, developing optimal pre-harvest strategies in these animals to control entry of non-O157 STEC into the food chain is as critical as it is for O157 (Griffin, 1995; Kaper et al., 1998; Griffin et al., 2000; Gyles 2006). In this context, understanding host–pathogen interactions is essential to the development of such strategies. We recently demonstrated that O157 colonization of the recto-anal junction (RAJ) of cattle not only involves bacterial microcolony formation on the lymphoid follicle-associated columnar epithelial cells but also involves diffuse adherence to the adjacent RAJ squamous epithelial (RSE) cells (Kudva et al., 2011). We subsequently developed an adherence assay using RSE cells that was able to successfully differentiate between O157 and other nonpathogenic E. coli strains as well as between strongly adherent and moderately adherent O157 strains from various sources (Kudva et al., 2011; Kudva et al., 2012). The RSE adherence assay was also extrapolated to differentiate between Shigella serotypes (Kudva, 2012).
Since the non-O157 STEC serotypes are closely related to O157 (Kaper et al., 1998; Gyles, 2006), we hypothesized that these human pathogens would also interact with bovine gastrointestinal cells in a similar manner, and that studies ascertaining such interactions would likely provide insights into the colonization dynamics of these STEC in their ruminant reservoirs. Hence, we (1) evaluated interactions of the non-O157 STEC serotypes with the RSE cells using the RSE cell adherence assay; (2) determined the role of Intimin, a well-characterized, primary STEC adhesin, and other proteins encoded along with Intimin on the locus of enterocyte effacement (LEE) in adherence of non-O157 STEC to RSE cells via adherence-inhibition assays; and (3) evaluated the ability of hyperimmune sera generated in cattle against O157 to inhibit adherence of non-O157 STEC to RSE cells to determine cross-protection across the STEC serotypes. Our results demonstrate that the pattern of adherence of non-O157 to bovine RSE cells is similar to that of O157; however, non-O157 STEC appear to utilize RSE cell adherence mechanisms that are distinct from those employed by O157 to bind the same cells at the primary site of colonization in the bovine gastrointestinal tract.
Materials and Methods
Bacterial strains and sera
Bacteria
STEC serotypes used in this study are listed in Table 1. Both clinical and environmental strains were included for each serotype except O121 due to lack of availability. For each non-O157 STEC serotype, strains A and B were used as test strains and strains C were used as the “verification strain,” to verify the adherence patterns observed (Table 1). O157 strain EDL 933 (Perna et al., 2001) was included for comparative purposes.
Number, numbers assigned to the non-O157 STEC strains by the source institutions; U, unknown/untypeable; NADC, National Animal Disease Center/United States Department of Agriculture, Ames, IA; HUS, hemolytic uremic syndrome; HC, hemorrhagic colitis; STEC Center, The Thomas S. Whittam Shiga-toxin producing Escherichia coli Center, East Lansing, MI; ATCC, American Type Culture Collection, Manassas, VA.
Pooled antisera
Rabbit antisera targeting the recombinant LEE-encoded proteins, Tir, EspA, and EspB, and Intimin (National Animal Disease Center Stocks, NADC, Ames, IA) were combined together to prepare the pooled antisera suspension (Kudva et al., 2012) for testing at 1:5 to 1:100 dilutions. The specificity of each fraction comprising the pooled antisera was confirmed by reacting against both O157 cell lysates and the cognate protein in Western blotting experiments (data not shown). Rabbit sera (Sigma-Aldrich, St. Louis, MO) from healthy animals (normal rabbit sera), at a 1:5 dilution, were used as a control in each experiment.
Pooled hyperimmune bovine sera
Hyperimmune bovine sera was generated by challenging cattle with oral doses containing a mixture of O157 strains of bovine and human origin, including strain EDL 933 (Rice et al., 2003). For use in this study, sera collected from nine cattle that had shed O157 in feces for up to 2 months in that study were pooled together to constitute the pooled hyperimmune bovine sera (Rice et al., 2003). Studies of the quality of the pooled hyperimmune bovine sera using dot immunoblotting and Western blotting revealed robust immunoreactivity against a variety of O157 components, including O157 lipopolysaccharide, previously identified LEE-encoded proteins, and several novel proteins that have been identified by us in our ongoing studies (Kudva et al., 2006; Kudva et al., 2010; Kudva et al., unpublished data). Sera from healthy cattle not challenged with O157 and being used as “blood donors” at the NADC, Ames, IA (normal bovine sera) were used as control at a 1:5 dilution, in each experiment.
RSE cell assays
RSE cells were obtained from the recto-anal junctions of the gastrointestinal tracts of healthy cattle as described previously (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012) and used in the following assays.
Adherence assays and patterns
Adherence of O157 to the RSE cells was previously demonstrated and developed into an adherence assay in our laboratory (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012). In this study, the assay was used to study the interactions of non-O157 STEC with RSE cells. Each of the RSE adherence assays was conducted in eight technical and two biological replicates as described previously (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012) for the test strains, and as eight technical replicates for the verification strain. Briefly, RSE cells were washed and resuspended in DMEM-No Glucose (Invitrogen, Carlsbad, CA) with 2.5% D+Mannose, to a final concentration of 105 cells/mL. Bacterial pellets from overnight cultures in DMEM-Low Glucose, incubated at 37°C without aeration, were washed and mixed with RSE cell suspensions to a final bacteria:cell ratio of 10:1 and incubated with aeration (37°C, 110 rpm, for 4 h). At the end of 4 h, the mixture was pelleted and washed thoroughly, once with 14 mL DMEM-NG and twice with 14 mL of sterile, distilled water (dH2O) before reconstituting in 100 μL dH2O. Eight 2-μL drops of this suspension were placed on Polysine (Thermo Scientific Pierce, Logan, UT) slides and dried overnight under direct light to quench nonspecific fluorescence, before fixing in cold 95% ethanol for 10 min. The slides were then stained with 1% toluidine blue, or with fluorescence-tagged antibodies that specifically target O157 or E. coli (Kierkegaard Perry Laboratories, Gaithersburg, MD) and the RSE cell cytokeratins as described previously (Kudva et al., 2011). Each experiment was then done in duplicate.
O157 adherence patterns on RSE cells were recorded as either diffuse or aggregative (clumps) for all positive interactions that involved direct association with the cells, as described previously (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012). Bacteria not adhering to cell membranes were considered to be negative for adherence (nonadherent) to the epithelial cells (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012). For a total of 100–160 well-dispersed RSE cells (10–20 cells per drop or chamber) analyzed per slide, the percent RSE cells with and without bacteria adhering to them were determined, and the average percents with standard error of means between trials was calculated using the GraphPad Prism5 software (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012). If more than 50% of RSE cells had >10 bacteria attached, the adherence was recorded as strongly positive. For >50% RSE cells with 1–10 adherent bacteria, the adherence was recorded as moderately positive. For less than 50% RSE cells with 1–5 adherent bacteria, the result was recorded as nonadherent.
Adherence-inhibition assay
The ability of pooled rabbit polyclonal antisera and bovine hyperimmune sera to interfere with and inhibit STEC adherence to RSE cells was evaluated. The assay was performed as before except that the bacterial cells were resuspended in sterile saline with or without antisera (“no sera” control), and incubated at 37°C for 30 min before mixing with RSE cells (Kudva et al., 2012). Both sera were tested at dilutions ranging from 1:5 to 1:100. Adherence patterns were recorded as described above.
HEp-2 cell adherence and adherence-inhibition assay
The HEp-2 cell adherence assays were performed with non-O157 STEC as described previously with O157 (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012) and under conditions used for RSE cell assays above. Each assay was conducted in duplicate, and in six chambers (technical replicates) of the chamber slides per run. Slides were stained with fluorescence-tagged antibodies that specifically target O157 or E. coli (KPL, Gaithersburg, MD), and the HEp-2 cell actin filaments as described previously (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012) and adherence patterns recorded as for RSE cells (see above).
Results and Discussion
Non-O157 STEC that were not exposed to any sera, irrespective of their serotypes or clinical/environmental derivation, adhered to RSE cells as well as the HEp-2 cells (Figs. 1A and 2A). Interestingly, the non-O157 STEC adherence patterns were similar to that of O157 strains as seen in this (O157 strain EDL 933) and previous studies (O157 strain 86-24) (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012). Of the seven non-O157 STEC serotypes tested, O145 and O111 showed a distinct aggregative binding pattern, while the other serotypes demonstrated diffuse adherence to RSE cells, similar to O157 strains EDL 933 and 86-24, respectively. However, all STEC adhered to HEp-2 cells in a diffuse manner. The differential adherence patterns are shown in Figs. 1A and 2A.
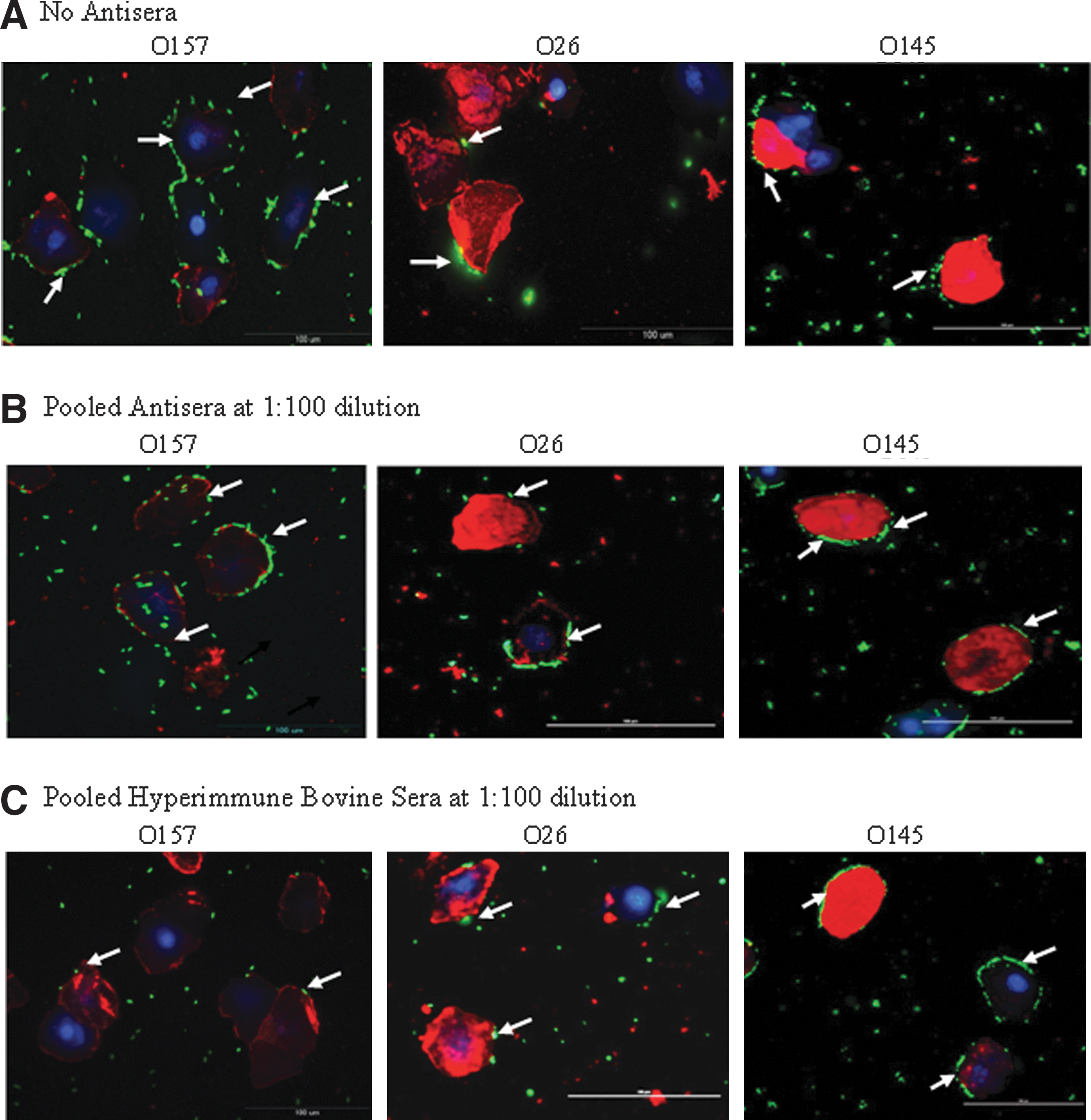
The different Shiga toxin–producing Escherichia coli adherence patterns, as represented by serotypes O157, O26, and O145, on recto-anal junction squamous epithelial cells in the presence of D+Mannose, and in the presence or absence of sera are shown.
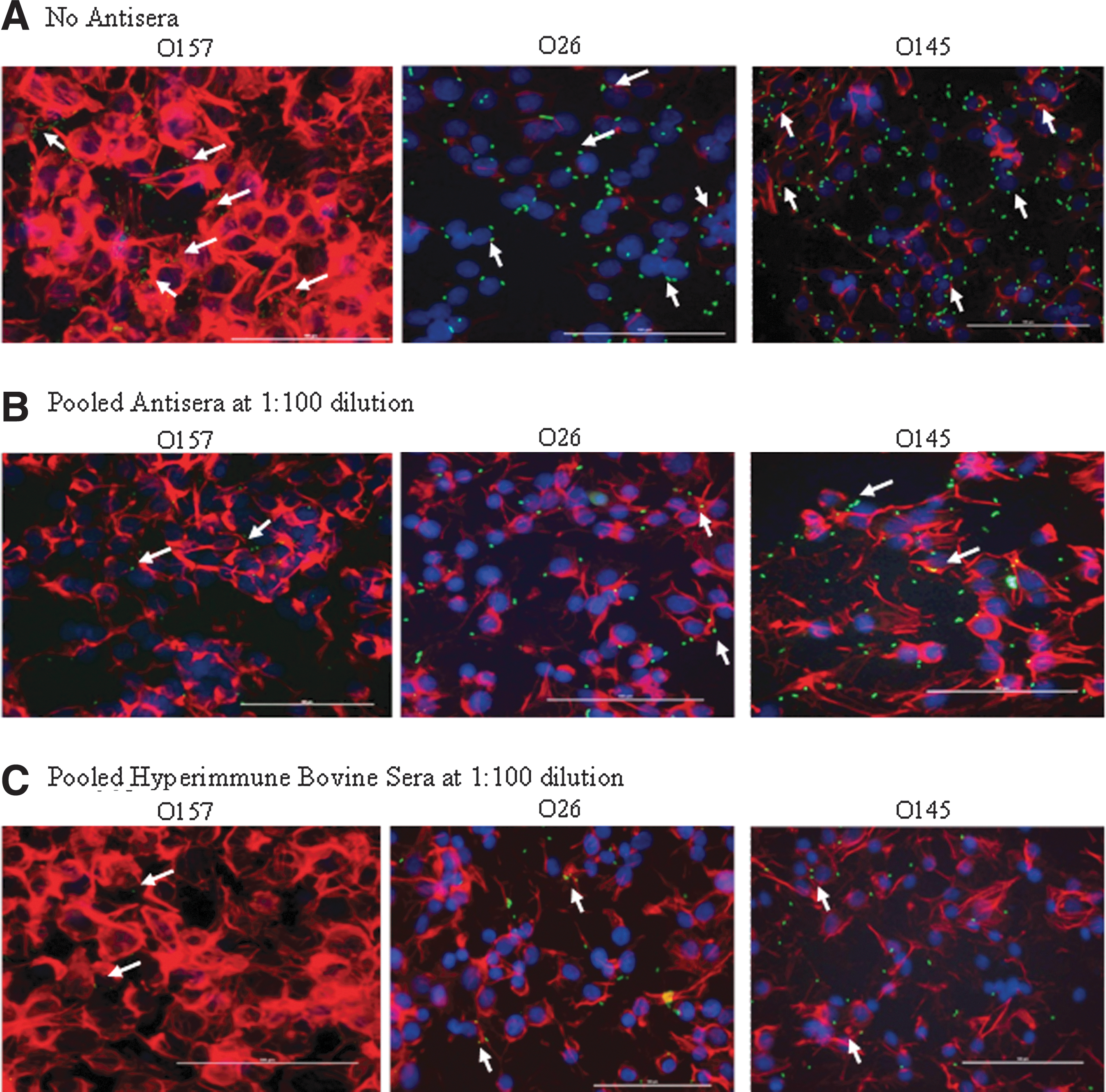
The different Shiga toxin–producing Escherichia coli adherence patterns, as represented by serotypes O157, O26, and O145, on HEp-2 cells in the presence of D+Mannose, and in the absence or presence of sera are shown.
As seen with O157 (Kudva et al., 2012), Intimin and other LEE-encoded proteins did not play a role in non-O157 STEC adherence to the RSE cells (Table 2 and Fig. 1B). The LEE-encoded proteins, which include EspA, EspB, Tir, and Intimin, have been shown to be required for efficient O157 adherence to the follicle-associated epithelium at the RAJ (Naylor et al., 2003; Naylor et al., 2005). However, in our recent studies, we demonstrated that these proteins were not involved in O157 adherence to the RSE cells also found at the RAJ (Kudva et al., 2012). Likewise, in this study, we found the non-O157 STEC adhered to RSE cells in the presence of pooled antisera containing antibodies blocking the Intimin, EspA, EspB, and Tir proteins (Table 2 and Fig. 1B). This was observed at all antisera dilutions tested. In fact, O113, O145, and O111 serotypes adhered strongly and in aggregative patterns in the presence of this pooled antisera, which may be indicative of other adhesins being better exposed by the masking of Intimin and the LEE proteins by this antisera (Kudva et al., 2012). Adherence to the RSE cells was also observed irrespective of the type of Intimin expressed (β-Intimin in serotype O45) (Oswald et al., 2000), or the lack of Intimin expression in some of the O113, O103, O121, O45, and O145 strains tested (Table 1), supporting a role of additional adhesins (Kudva et al., 2012).
Average of two trials conducted for 2 strains, A and B, of each serotype, except for serotype O121:H19 for which only one strain was available, is shown. Each trial had one slide per bacterial group. Each slide in turn had eight technical replicates spotted on it; 10–20 well-dispersed RSE cells were evaluated per spot. Percent means for ranges used to determine “moderate or strong” adherence, in the absence of any antisera, are shown in bold.
Number of bacteria adhering to each RSE cell is shown as a range of >10, and 1–10. Number of RSE cells without bacteria is not shown.
STEC, Shiga toxin–producing Escherichia coli; MOI, multiplicity of infection.
Surprisingly, despite similarities in RSE adherence, the pooled hyperimmune bovine sera had opposite effects on the various STEC serotypes. A cross-inhibitory effect was anticipated with the pooled hyperimmune bovine sera that were generated in cattle experimentally inoculated with multiple strains of O157, considering the existing similarities between the STEC (Kaper et al., 1998; Gyles, 2006). However, these sera demonstrated serotype specific inhibition by blocking only the adherence of O157 to RSE cells and none of the non-O157 STEC (Table 2 and Fig. 1C), at all dilutions tested.
The HEp-2 adherence assays are routinely used to evaluate adherence potential of O157 and non-O157 STEC (Kaper et al., 1998; Vidal et al., 2008), and have provided useful information in the context of the human host. Although the inability of these assays to reflect O157 interactions with the bovine RSE cells was already established (Kudva et al., 2011; Kudva et al., 2012), we included the HEp-2 cell adherence assay to compare results obtained with non-O157 STEC and the RSE cells. No differences were observed in the inhibitory effects of the pooled antisera or hyperimmune bovine sera on the STEC adherence to HEp-2 cells, irrespective of the serotype and sera dilutions used. Sera generated against O157 blocked adherence of various STEC serotypes to HEp-2 cells, which was in accordance with results reported by other researchers as well (Asper et al., 2007; Moradi-Bihendi et al., 2010). In fact, all isolates were effectively inhibited from adhering to HEp2 cells by both the pooled antisera (Table 3 and Fig. 2B), and the hyperimmune bovine sera (Table 3 and Fig. 2C), which was in contrast to the observations made with the RSE cells (Table 2).
Average of two trials conducted for two strains, A and B, of each serotype, except for serotype O121:H19 for which only one strain was available, is shown. Each trial had one slide per bacterial group. Each slide in turn had six technical replicates in separate chambers on it; 10–20 well-dispersed HEp-2 cells were evaluated per chamber. Percent means for ranges used to determine “moderate or strong” adherence, in the absence of any antisera, are shown in bold.
Number of bacteria adhering to each HEp-2 cell is shown as a range of > 10, and 1–10. Number of HEp-2 cells without bacteria is not shown.
MOI, multiplicity of infection; STEC, Shiga toxin–producing Escherichia coli.
Non-O157 STEC adhered to RSE cells as well as or better than O157, which may reflect upon the close relatedness of these STEC serotypes with O157 (Gyles, 2006). In this context, three significant observations were made in this study, including the following: (1) Intimin and other LEE-encoded proteins are not involved in the adherence of non-O157 STEC to RSE cells as reported for O157 (Kudva et al.,), (2) hyperimmune cattle sera prevents adherence of homologous but not heterologous STEC serotypes to RSE cells, and (3) STEC interactions with cells derived from natural reservoir/animal hosts are not accurately reflected by those seen with human-derived HEp-2 cells as previously reported (Kudva et al., 2011; Kudva, 2012; Kudva et al., 2012), which dictates exercising caution in extrapolating inferences from results obtained using human cell-based assays when it comes to developing host-specific management modalities.
The central of the three observations made in this study (namely, that a natural O157 infection [or suboptimal experimental inoculation] does not elicit immune responses capable of abrogating adherence of heterologous non-O157 STEC to bovine RSE cells) has important implications for development of efficacious modalities, including broadly protective STEC vaccines. It has been well documented that constituents of such vaccines, with highly conserved proteins and domains that include conserved epitopes, are subject to pathogen decoy mechanisms because they serve important functions to the pathogen (Harty et al., 1988; Embers et al., 2002; Crane et al., 2006; Bianchi et al., 2005; Gambhira et al., 2007; Sollner et al., 2008; Alphs et al., 2008; Chootong et al., 2010). The end result of exertion of such decoy mechanisms is that highly conserved proteins/epitopes are rendered subimmunodominant during natural infection or suboptimal vaccination (Crane et al., 2006). Hence, such highly conserved proteins/epitopes targeted by host immune responses (Corti et al., 2011) engender only subthreshold antibody responses that are ineffective in mediating heterologous protection. Interestingly, however, when such conserved antigens are identified and delivered optimally to the immune system (as in an efficacious vaccine), they reportedly confer excellent cross-protection (Crane et al., 2006). Therefore, the obvious challenge is to identify such protein components of broadly protective STEC vaccines. We are currently addressing this challenge by adopting innovative strategies, including systems-based approaches to dissect alternate mechanisms of adherence used by these STEC to bind RSE cells with the objective of identifying novel shared adhesins.
Footnotes
Acknowledgments
Technical assistance provided by Bryan Wheeler, Denise Chapman, and the animal caretakers at the NADC, Ames, IA is acknowledged. The work of C.J.H. was supported, in part, by the Public Health Service NIH grants P20-RR16454 (NCRR) and P20-GM103408 (NIGMS). We especially acknowledge Dr. Terrance Arthur, Clay Center, NE and Dr. Brian Brunelle, NADC, Ames, IA for their insightful review of this manuscript.
Disclosure Statement
No competing financial interests exist.