
Abstract
The survival of Salmonella enterica serovar Thompson and Escherichia coli O157 was investigated on growing butterhead lettuce plants in the plant-growth chamber and greenhouse. All inoculation tests were made under conditions that approximate the greenhouse conditions for butterhead lettuce cultivation in Flanders (Belgium). The survival and proliferation of the pathogens on the leaves was determined at days 0, 4, and 8 after inoculation using standard plating techniques on selective medium. In the growth chamber, the extent to which both pathogens were able to multiply on the lettuce leaves was influenced by crop stage and leaf age. On young plants, the older leaves supported pathogen survival better. On nearly mature plants, pathogen population sizes were significantly higher on the old and young leaves compared with middle-aged leaves (p<0.001). In the greenhouse, the environmental regimen with high fluctuations in temperature and relative humidity was less conducive to the survival of E. coli O157, though its survival on nearly mature lettuce was enhanced by overhead irrigation. The moist conditions between the folded inner leaves are likely contributing to the survival of enteric pathogens in the lettuce head. Butterhead lettuce grown in greenhouses with a sprinkle irrigation system may present a potential health hazard when contaminated near harvest. Experimental design (growth chamber versus greenhouse) largely influences enteric pathogen behavior on growing lettuce plants.
Introduction
G
Plant surfaces may become contaminated before harvest. Irrigation water is considered a potentially important introduction route, and several studies have shown its potential for the transmission of enteric pathogens to lettuce plants. However, most of these studies were conducted either under laboratory or controlled conditions in growth chambers (Brandl and Mandrell, 2002; Solomon et al., 2003; Aruscavage et al., 2008; Aruscavage et al., 2010; Brandl and Amundson, 2008; Zhang et al., 2009; Erickson et al., 2010a) or field experiments (Barker-Reid et al., 2009; Erickson et al., 2010b; Harapas et al., 2010; Wood et al., 2010; Fonseca et al., 2011). Few studies have compared both types of experimental setups. Furthermore, these experiments were often performed with lettuce cultivars and environmental conditions typical for the production of leafy vegetables in the United States.
This study focused on butterhead lettuce (Lactuca sativa L. var. capitata), an important leafy vegetable grown commercially in Northern European countries and regions such as Flanders (Belgium) mainly for export. The crop is cultivated in greenhouses in a continuous monoculture system using an overhead sprinkler irrigation (Pauwelyn et al., 2011), creating conditions that, in case of contaminated irrigation water, are likely to deposit enteric pathogens on the lettuce plants. No scientific information is available concerning enteric pathogens for this lettuce cultivar in combination with its commercial growing conditions. The aim of this study was to investigate the effects of leaf age (young, middle-aged, and old), crop stage (before or after heading), and daily overhead irrigation on the survival of E. coli O157:H7 and Salmonella enterica should they be introduced by irrigation water on butterhead lettuce in Northern European greenhouses.
Materials and Methods
Strains and growth conditions
Salmonella Thompson RM1987N is a spontaneous nalidixic-acid-resistant mutant of a described clinical isolate from a patient in a cilantro-linked outbreak (Brandl et al., 2005) and was kindly donated by Dr. Maria Brandl (USDA-ARS, Albany, CA). E. coli O157:H7 MB3885 naturally lacks Shiga toxin genes (stx1 and stx2) and was used as a nontoxigenic surrogate strain for the Shiga toxin–producing serotype O157:H7 for biosafety reasons. This strain originates from beef carpaccio and was kindly donated by the Scientific Institute for Public Health (Brussels, Belgium). The absence of stx1 and stx2 genes and presence of other virulence genes eae (intimin), ehx (enterohemolysin), espP (extracellular serine protease), and katP (catalase–peroxidase) were confirmed by conventional PCR (Verstraete et al., 2012). Both strains were streaked from a glycerol frozen stock maintained at −70
Plant growth conditions
Pelletized butterhead lettuce seeds (Lactuca sativa L. var. capitata “Alexandria”) were obtained from Rijk Zwaan Distribution B.V., De Lier, the Netherlands. The seeds were sown in ground blocks of 4×4×6 cm (seed and cutting compost, Saniflor, Geraardsbergen, Belgium). Two weeks after sowing, the seedlings were placed in pots of 13-cm (for the experiments with young plants, nine-leaf stage) or 20-cm diameter (for the experiments with nearly mature plants) and grown in the greenhouse at Institute for Agricultural and Fisheries Research (ILVO). For inoculations in the growth chamber, the plants were moved from the greenhouse to the growth chamber (Isocab, Harelbeke, Belgium) 2 days before inoculation and placed in trays with±2-cm irrigation water. Growth chamber conditions were set at ±19°C during the day and at ±12°C at night with a relative humidity (RH) of 70–80% and a photoperiod of 14 h. The greenhouse experiment was conducted in April 2010, and conditions of RH and temperature were logged in 5-min intervals using an EL-USB-2 data logger (Lascar Electronics, Salisbury, UK) (Table 1).
SD, standard deviation.
Plant inoculations and irrigation
Cells of each strain were washed twice by centrifugation (6000×g, 15 min) in 50 mM phosphate buffered saline (PBS, pH 7.4). The optical density (OD) was measured at 595 nm using a microplate reader (Bio-Rad 3550, Richmond, CA) and by using an OD–colony-forming unit (CFU)/mL standard curve, the appropriate amount of cells was resuspended in PBS to give 1×105 CFU/mL. For each combination of pathogen and crop stage, a total of 14 plants were inoculated. Young plants were inoculated by immersion as described by Brandl and Amundson (2008). Nearly mature plants were spray-inoculated as immersion was not feasible for biosafety reasons. The leaves were sprayed with a hand-held sprayer until runoff, which corresponded with approximately 100 mL inoculum per plant. A preliminary test was performed to compare both inoculation methods. This test showed similar survival trends of the pathogens. The pots were wrapped with plastic film (Saran Wrap; Dow Chemical Company) to prevent soil contamination by dripping. Both inoculation methods resulted in a pathogen level of approximately 3×103–104 CFU per gram of leaf tissue as determined by selective plating as described hereafter. The control treatment (six plants) consisted of PBS without added inoculum. To test the influence of overhead irrigation, half of the plants in each experiment were also watered from above with a hand-held sprayer until runoff, whereas the other plants were only watered from below. For all experiments, irrigation water from the experimental greenhouse at ILVO was used, which originates from ground water. A chemical analysis of the water was performed by Inagro (Rumbeke-Beitem, Belgium) and ILVO (Merelbeke, Belgium) (Table 2).
EC, electrical conductivity; BOD, biological oxygen demand; COD, chemical oxygen demand.
Measurement of pathogen populations on lettuce leaves
Before inoculation, the leaves of young plants were labeled from old to young with small pieces of tape. Old, middle-aged, and young leaves were numbered 1–3, 4–6, and 7–9, respectively. For inoculated nearly mature lettuce heads, the fourth to sixth leaves were sampled as old leaves, the 12th to 14th leaves as middle-aged, and the leaves enclosed in the head as young leaves. Three randomly chosen plants from each treatment (i.e., pathogen–crop stage–irrigation combination) were sampled at 4 and 8 days after inoculation, while one plant per treatment was sampled at day 0. From each plant, three leaves from each leaf age category were collected using sterile instruments. The leaves were cut approximately 1 cm above the soil surface, placed individually in a sterile extraction bag with filter (Bioreba, Reinach, Switzerland), and each leaf was weighed. Then, PBS with 0.05% vol/vol Tween 20 was added at a 1/1 (wt/vol) ratio and the sample was ground for ±15 s at maximum speed (Homex 6, Bioreba) until a homogenous mixture was obtained. Tenfold dilutions of the resulting suspension were made in 0.1% peptone and spread-plated on xylose lysine desoxycholate (XLD, LAB032; Lab M, Bury, UK) overlaid with TSA for Salmonella (XLD-TAL) or cefixime–tellurite sorbitol Mac Conkey (CT-SMAC, Lab 161; Lab M, Bury, UK) overlaid with TSA (CT-SMAC-TAL). The TAL method was described earlier for the resuscitation of injured cells (Kang and Fung, 2000; Qiu and Wu, 2007). All plates were incubated at 37°C for at least 18 h. Simultaneously, a nonselective enrichment was conducted by adding 1 mL of leaf homogenate to 9 mL of buffered peptone water (Oxoid) and incubated at 37°C, 200 rpm for 24 h. Only when the plate counts were below the detection limit (<1.3 log CFU/g), 100 μL of the enrichment culture was streaked onto the appropriate selective medium.
Two different controls were performed to ensure that the inoculated strains and not the indigenous bacteria were counted. First, the undiluted leaf homogenate of the control plants was plated onto XLD-TAL and/or CT-SMAC-TAL. Second, randomly selected and presumptive nonpathogen colonies were subjected to a serological test (E. coli O157:H7: DR0620, Oxoid, Basingstoke; Salmonella: DR1108, Oxoid, Basingstoke).
Statistical analyses
The data were analyzed in SPSS (IBM SPSS Statistics 19) using a multi-factor analysis of variance (linear models) with α=0.05. The data were normalized by subtracting the mean of the log count at day 0 from the mean of the log-transformed values at days 4 and 8. The explanatory fixed variables were day (day 4 or day 8), leaf age (young–middle-aged–old) and overhead irrigation treatment (with or without). Full factorial design was first performed. If all the interaction terms were not significant, a simplified model without interaction could be used with the Scheffé-test as post-hoc test. In other instances, Fisher's Least Significant Difference post-hoc comparisons were made with the combinations of the groups. Values below the detection limit were considered to be at the detection limit (1.3 log CFU/g) if the enrichment was positive, whereas those testing negative by enrichment culture were assigned a value of 0.0 as described by Erickson et al. (2010a).
Results
Survival and growth potential on young plants in the growth chamber
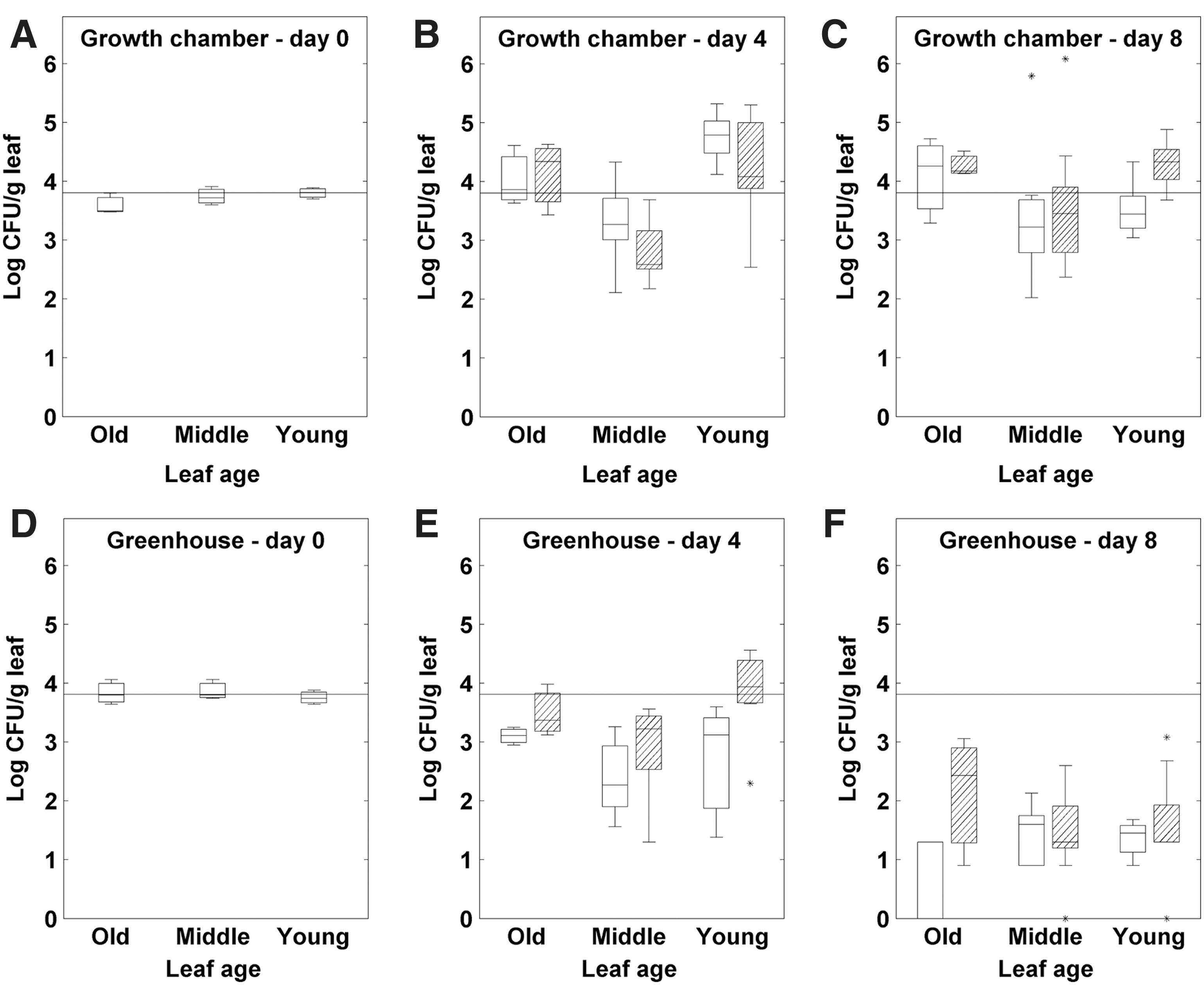
The results are shown in Figure 1. Both pathogens were able to proliferate on lettuce leaves after inoculation. For Salmonella, no significant differences were found between the average counts at 4 and 8 days after inoculation (p>0.05), whereas for E. coli O157 a slight increase could be observed (0.01<p<0.05). For both pathogens, no significant differences were identified between plants with and without daily overhead irrigation, and survival on the youngest leaves was significantly lower than on the middle-aged or oldest leaves (p<0.001). For Salmonella (Fig. 1A–C), the viable counts decreased by 1.14 log CFU/g on the young leaves, whereas they slightly increased on the middle-aged and old leaves by 0.38 and 0.54 log CFU/g, respectively (average of days 4 and 8). For E. coli O157 (Fig. 1D–F), the same trend was observed but with a greater relative increase. On average, the E. coli O157 counts on the young leaves increased slightly (0.40 log CFU/g) and showed a greater increase on the middle-aged and old leaves (1.86 and 1.99 log CFU/g, respectively). At day 8, however, this leaf age effect was no longer measurable for the daily irrigated plants inoculated with E. coli O157 (Fig. 1F). Generally, the pathogen population sizes greatly varied from one leaf to another, even between leaves of the same leaf age class.
Growth chamber experiment with young butterhead lettuce (nine-leaf stage): pathogen population dynamics on young, middle-aged, and old leaves at 0, 4, and 8 days after inoculation.
Survival and growth potential on nearly mature plants in the growth chamber
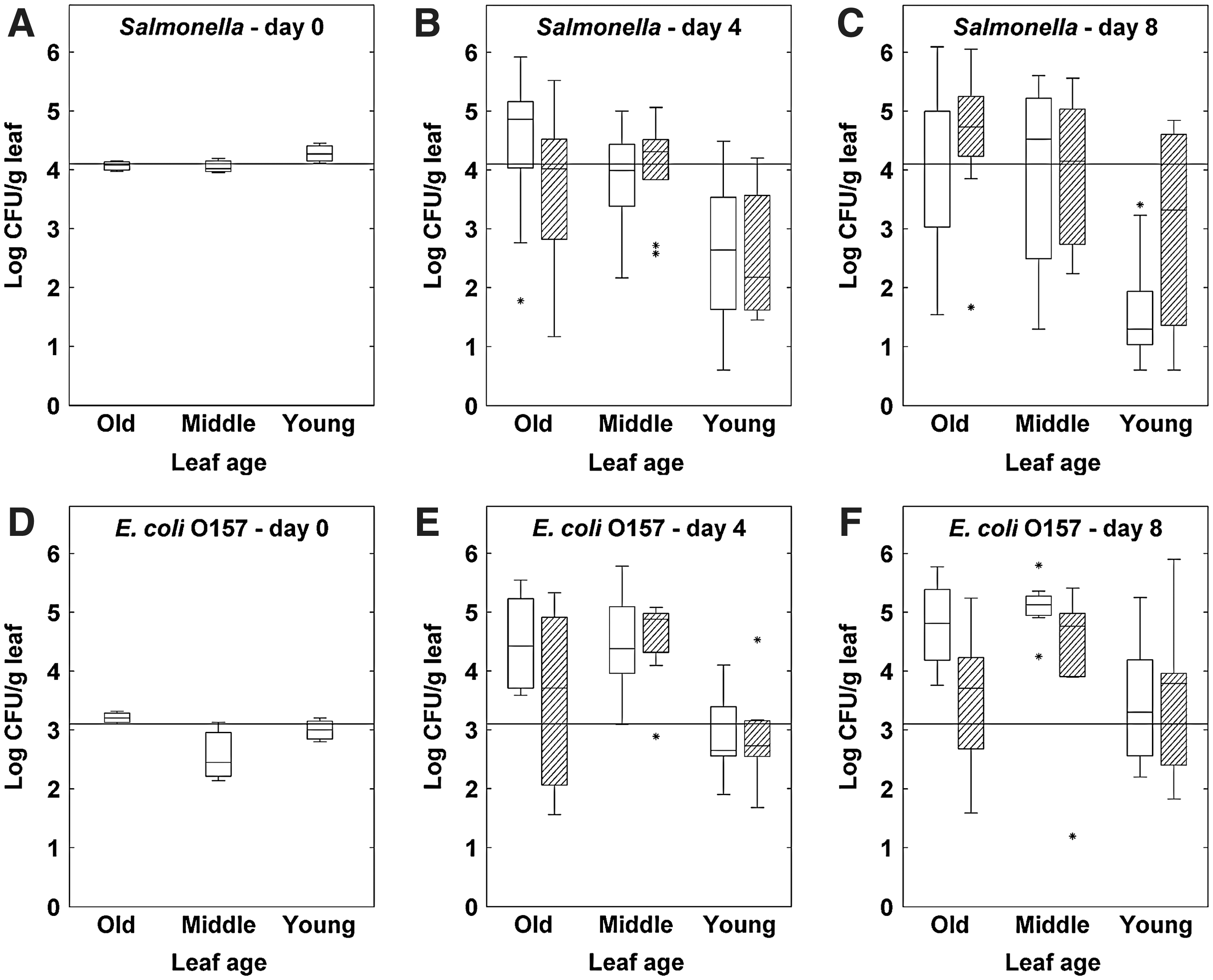
The results are shown in Figure 2A–C. Leaf age had a significant effect (p<0.001) and overhead irrigation had no effect. Leaf age did not have the same effect for nearly mature plants as compared to the young plants: E. coli O157 proliferated on young inner leaves and oldest outer leaves (an average increase of 0.67 and 0.28 log CFU/g, respectively) and decreased on the middle-aged lettuce leaves (on average−0.76 log CFU/g). This leaf age effect was most obvious on day 4 and was no longer observed on day 8 after inoculation. On average, the total number of pathogens per plant did not increase significantly during the 8 days in the growth chamber. Similar trends were also observed for Salmonella, though this experiment was only conducted until day 4 (see Supplementary Fig. S1) (Supplementary Data are available online at
Preharvest population dynamics of Escherichia coli O157:H7 strain MB3885 on young, middle-aged, and old leaves of nearly mature butterhead lettuce plants at 0, 4, and 8 days after inoculation in the growth chamber and in the greenhouse.
Survival and growth potential on nearly mature lettuce in the greenhouse
The results are shown in Figure 2D–F. On average, no increase in pathogen counts was observed, and the survival of E. coli O157 was significantly lower than in the experiment conducted in the growth chamber. On day 4, E. coli O157 survived the best (initial inoculum density was retained) on young, overhead-irrigated leaves (p<0.01). Only at day 4, a leaf age effect existed between young and middle-aged leaves of the overhead-irrigated plants. On day 8, the E. coli O157 level was significantly higher on old leaves of overhead-irrigated plants, whereas an enrichment step was needed to detect the pathogen on old leaves of nonirrigated heads (0.01<p<0.05). On the middle-aged leaves, no statistically significant influence of overhead irrigation was observed. The average number of E. coli O157 had decreased to 1.54±0.88 log CFU/g for the daily overhead-irrigated plants, and to 1.38±0.38 log CFU/g leaf for the nonirrigated plants. In total, 13 of the 54 samples had to be enriched but in 10 of these 13 samples, the pathogen could still be detected. This experiment could not be performed for Salmonella for biosafety reasons.
Discussion
In the present study, we looked at plant-related factors such as leaf age, crop stage, and overhead irrigation to see whether these can have a potential influence on the survival of enteric pathogens in the phyllosphere of lettuce. In comparison with other studies on the survival of these pathogens on fresh produce (e.g., Aruscavage et al., 2008; Brandl and Amundson, 2008; Erickson et al., 2010a), we used growing butterhead lettuce plants cultivated under realistic northern European lettuce production conditions. The experiments were performed under the controlled conditions of a growth chamber, but in parallel an experiment in the greenhouse was made to evaluate whether the investigated factors are also important under conditions of current practice.
The experiments in the growth chamber revealed that lettuce leaf age in combination with crop stage has an effect on the survival capacity of the pathogen. The first evidence for this phenomenon was presented by Brandl and Amundson (2008). The experimental design of our growth chamber study was quite similar to theirs, but differed in the used lettuce cultivar (butterhead lettuce with softer folded leaves and closed head formation versus romaine lettuce with upright long leaves and semi-open head formation), lower temperatures (19°C day–12°C night versus constantly 28°C), lower RH (70–80% versus 100%) and longer sampling period (8 days versus 2–3 days). For nearly mature plants, the pathogen population dynamics on the leaves at day 4 after inoculation were in agreement with their findings. The highest population size of E. coli O157 found on the young inner leaves, which are preferably consumed, was on average 1.4 log CFU/g higher than on the middle-aged leaves. However, at day 8 after inoculation, when the plants were ready for harvest, this young-leaf effect was no longer observed. Brandl and Amundson (2008) reported a similar young-leaf effect for young plants as well as for nearly mature plants. In contrast, the population size of the pathogens on young plants in our study was on average 1.60 log CFU/g lower compared to the old and middle-aged leaves. Our results do not necessarily contradict the findings of Brandl and Amundson (2008), as we may have missed the young-leaf effect on young plants because of our later sampling of day 4 after inoculation. This was also suggested by an additional experiment that included a sampling at day 1 after inoculation whereby growth of the pathogen was noted after 1 day (data not shown). Several factors may explain our different results on young leaves of young plants. During the 8 days in the growth chamber, the young leaves became more mature and the nutrient availability may have changed. Brandl and Amundson ascribed the leaf-age-dependent growth to the different nitrogen content in the exudates from leaves of different ages. The bigger relative change in leaf size of young leaves of young plants may also have an effect on the pathogen counts. As an illustration, the average weight of the young leaves increased by ±36%, whereas the old and middle-aged leaves of the young plants did not grow much. Also, it is likely that in contrast to the folded young leaves of nearly mature lettuce, the younger leaves of young plants are more susceptible to desiccation over time because they lack protection from other leaves.
Several authors have highlighted the importance of the RH conditions for survival of bacterial pathogens on plants (O'Brien and Lindow, 1989; Brandl and Mandrell, 2002; Stine et al., 2005). It was shown that Salmonella could barely grow on plants that were kept below 40–50% RH but that short periods of high RH were sufficient to recover maximum population size on the leaves (Brandl and Mandrell, 2002). Therefore, we expected that daily overhead irrigation, hence higher humidity, could lead to an increased growth of the pathogens on the leaves. However, in the growth chamber additional daily overhead irrigation on top of the relative high humidity (70–80%) had no effect on the behavior of the pathogens. In the greenhouse, the effect of overhead irrigation was only sporadic. The RH was 45% on average during the day with higher values up to 76% measured at night, but this did not seem to be sufficient for resuscitation of E. coli O157 on the leaves. The plants were irrigated daily in the morning, but after 2 h most of the water on the leaves was already evaporated; therefore, the period of higher relative humidity may not have been long enough. If pathogens are internalized into the leaf tissue, the irrigation treatment may have no effect as well. This could not be investigated because the set-up used for our study did not allow for making a distinction between endo- and epiphytical pathogen populations.
Furthermore, large fluctuations in temperature were measured in the greenhouse. The influence of such fluctuations has not been intensively studied. For natural substrates, Semenov et al. (2007) showed that E. coli O157 and Salmonella inoculated in cow manure were very sensitive to fluctuating temperatures. Future research should be directed to investigate the role of environmental fluctuations on plant–pathogen interactions in order to gain better insights into the fitness of enteric pathogens on plants grown under commercial conditions.
Conclusions
In this study, it was demonstrated that leaf age, crop stage, and experimental design can have an important influence on the survival and proliferation of E. coli O157 and Salmonella on butterhead lettuce. Greenhouse conditions with high fluctuations in temperature and RH were less favorable for enteric pathogen growth compared to controlled conditions. The impact of daily overhead irrigation was only observed in the greenhouse on nearly mature plants, indicating that sprinkler irrigation may increase food safety risk, especially after lettuce heading.
Footnotes
Acknowledgments
We wish to thank Dr. Maria Brandl and the Scientific Institute for Public Health (Brussels, Belgium) for providing strains, Marleen Seynnaeve and Chris Van Waes for analyzing the irrigation water sample, and Cinzia Van Malderghem for excellent technical support. This study was funded by the Federal Public Service of Health, Food Chain Safety and Environment (contract SALCOSLA RF 6202).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.