
Abstract
Poultry and poultry products are major contributors of zoonotic pathogens. Limited data are available on Enterobacter spp. as a potent zoonotic pathogen in poultry. The present study is a first endeavor on the emergence of multidrug-resistant zoonotic Enterobacter spp. and its prevalence arising from poultry in Bangladesh. Cloacal swabs from poultry samples of five different farms at Savar, Dhaka, Bangladesh were collected and from 106 isolates, 18 presumptive Enterobacter spp. were obtained. Antibiogram using 19 used antibiotics belonging to 15 major groups revealed that all of the 18 isolates were completely resistant to penicillin and rifampicin, but differed in their drug resistance pattern against ampicillin (94.4%), clindamycin (94.4%), erythromycin (94.4%), vancomycin (88.9%), sulfonamides (72.2%), imipenem (66.6%), streptomycin (55.6%), nitrofurantoin (33.3%), doxycycline (33.3%), tetracyclines (33.3%), cefepime (11.1%), and gentamicin (5.6%). All Enterobacter spp. were found to be plasmid free, implying that multidrug-resistant properties are chromosomal borne. The vanA and sulI were detected by polymerase chain reaction assay in 17 and 13 isolates, respectively. Amplified ribosomal DNA restriction analysis and randomly amplified polymorphic DNA distributed the 18 multidrug-resistant Enterobacter spp. into three genotypes. Phylogenetic analysis of the representatives of the three genotypes using partial 16S rRNA gene sequence (approximately 900 bp) showed that the genotypically diverse groups belonged to Enterobacter hormaechei, E. cloacae, and E. cancerogenus, respectively. The clinical significance of the close relative Enterobacter spp. is indicative of their zoonotic potential. Therefore, urgent intervention is required to limit the emergence and spread of these bacteria in poultry feed as well as prudent use of antibiotics among poultry farmers in Bangladesh.
Introduction
R
The poultry industry, with 1.79 million poultry and livestock farms (data collected from Livestock economist section, Department of Livestock service, Dhaka), is a leading industry in Bangladesh, and about 3 million people are employed in this sector. This industry has immense potential for boosting the economic growth of the country as well as ensuring food security for protein sources. The scenario of antibiotic resistance in poultry of Bangladesh is currently a serious issue. Bulks of antibiotics including renamicin, ciprofloxacin, erythromycin, colistin, cephalosporin, etc. are used (either overused or misused) in poultry and livestock sectors of Bangladesh. Unfortunately, a very large portion of this is used under conditions of inadequate or no medical supervision and in most cases without prior tests on identification of the disease-causing organism and determination of its sensitivity to the antibiotic prescribed. Therefore, emergence of multidrug-resistant (MDR) zoonotic pathogens in poultry, particularly from feed, is alarming in Bangladesh (Faiz and Basher, 2011).
The genus Enterobacter is more specifically a nosocomial opportunistic pathogen and is sought as one of the many key causes for extraintestinal infections next to Escherichia coli (Brooks et al., 2007). Enterobacter species, particularly E. cloacae, E. hormaechei, and E. aerogenes, are important nosocomial pathogens responsible for lower respiratory tract infections, skin and soft-tissue infections, urinary tract infections, endocarditis, intra-abdominal infections, septic arthritis, osteomyelitis, central nervous system, and ophthalmic infections (Mahapatra et al., 2002; Manzur et al., 2007; Moriguchi et al., 2007; Dalben et al., 2008). Research investigations frequently reported antibiotic resistance of various Enterobacter spp., specifically E. cloacae, isolated from food animals such as ground beef cattle farm, processing facilities, and clinical settings (Kim and Wei, 2007). However, not much literature is currently available on the prevalence and antimicrobial resistance of Enterobacter in poultry and poultry products in the Asian region (Nazer et al., 2006), and no literature is currently available on the respective studies in Bangladesh. Considering the severity of developing antibiotic resistance and zoonotic potentials within Enterobacter spp., this study attempts to investigate the prevalence and characterizations of the circulating types of MDR zoonotic Enterobacter spp. from commercial poultry farms in Bangladesh.
Materials and Methods
Sample collection and isolation of Enterobacter spp.
Cloacal samples were collected from 106 birds of five poultry farms located in the semiurban area of Savar region of Dhaka, Bangladesh (Mahmud et al., 2011).
The Enterobacter-like suspected colonies from McConkey agar (Oxoid, UK) plates were picked up by sterile needle and re-streaked onto the respective plates as well as on Nutrient Agar (Oxoid) plates for further identification. Morphological characteristics of the colonies (shape, size, edge, elevation, form, and opacity) developed after 18–24 h of incubation on MacConkey agar media were recorded, and pure cultures were selected on the basis of microscopic observation and biochemical properties described in Bergey's Manual of Determinative Bacteriology (Bergey and John, 1994).
Antimicrobial susceptibility test
The antimicrobial susceptibility of the isolates was determined in vitro by using the standardized agar-disc-diffusion method (Bauer et al., 1966). Nineteen commercially available antibiotic discs (Oxoid) (see Supplementary Table S1) (Supplementary Data are available online at
Plasmid profile analysis
Plasmid DNA was extracted by using Wizard®Plus SV Minipreps plasmid DNA Purification kit (Promega, USA) and by manual extraction procedure (Birnboim and Doly, 1979). Plasmid DNA was analyzed by electrophoresis in 0.8% agarose gel. Plasmid extracted from E. coli V517 were used as control and for molecular weight determination.
Extraction and purification of chromosomal DNA
The chromosomal DNA was extracted from a fresh bacterial culture (<109 cells/mL) with ATP™ Genomic DNA Mini Kit (ATP Biotech Inc., USA) according to the kit manual.
Polymerase chain reaction
PCR was done with GoTaq® PCR Core System II (Promega, USA) using universal primers for bacterial 16S rRNA gene and primers specific for antibiotic resistance genes such as vanA, vanB, sulI, qnrA1, bla TEM, bla SHV, and bla CTX (see Supplementary Table S2) within a thermal cycler (Biometra, Germany). The PCR reaction was initial heating at 96°C for 5 min and then continued with the denaturation at 94°C, for 1 min 30 s, annealing at specified temperature (mentioned for each primer set in Supplementary Table S2), and extension at 72°C for 1 min 30 s for a total of 35 cycles followed by a final extension of 10 min at 72°C. PCR tubes were then stored at −20°C for analysis. The cycling profile for each primer:target combination was optimized accordingly.
Molecular fingerprinting of Enterobacter spp.
Amplified ribosomal DNA restriction analysis (ARDRA)
Complete digestion of 16S rRNA gene PCR product (approximately 900 bp) of the poultry Enterobacter isolates using the HaeIII (Promega, USA) restriction enzyme was done for ARDRA (Vaneechoutte et al., 1995). The reaction mixture contained 2 μL 10X buffer, 0.2 μL bovine serum albumin, 6U enzyme, 2.5 μL water, and ∼1000 ng PCR product. The digests were visualized after agarose gel electrophoresis, using 1.5% agarose (wt/vol) gel. Two different-size DNA markers, 1 Kb and 100 bp (Bioneer, South Korea) were used to analyze different restriction fragments.
Randomly amplified polymorphic DNA (RAPD) genotyping
In RAPD (Mbwana et al., 2006), two random primers (1283 and R2) (supplementary Table S2) were used for their ability to discriminate between species of Enterobacter isolates. PCR GoTaq® PCR Core System II kit (Promega, USA) conditions included initial denaturation at 95°C for 5 min, followed by 35 cycles of 1 min at 94°C, 1 min at 40°C, and 2 min at 72°C. Thirty-five cycles of these segments were repeated with a final extension of 10 min at 72°C. After amplification, the banding profiles were visualized under UV light and digitalized by the AlphaImager HP System Versatile Gel Imaging (USA).
Sequencing and phylogenetic analysis
The 16S rRNA gene PCR products of representative strains of Enterobacter spp. from each ARDRA and RAPD groups were purified with the Wizard PCR SV Gel and PCR Clean-Up System kit (Promega, USA) and sequenced (ABI Prism 3130 Genetic Analyzer, USA) using forwards 27F and reverse 907R primers (supplementary Table S2). Partial sequences were combined to full-length sequences (800 bp–900 bp) via the SeqMan Genome Assembler (DNAstar, USA) and were compared to the GenBank database of the National Center for Biotechnology Information (
Multiple sequence alignment of the retrieved reference sequences from National Center for Biotechnology Information was performed with the ClustalW (Larkin et al., 2007) software and was exported to the Molecular Evolutionary Genetics Analysis (MEGA) program (Tamura et al., 2007) for phylogenetic tree construction using the Neighbor joining algorithm and selecting 1000 bootstrap replication.
Nucleotide sequence accession numbers
The 16S rRNA gene sequences of the Enterobacter isolates have been deposited in the GenBank database (accession numbers JX872223- JX872227).
Results
Isolation and Identification of Enterobacter spp.
On the basis of colony morphology, 18 presumptive Enterobacter spp. from 106 isolates on the MacConkey Agar plates were grouped into three morphotypes (Fig. 1). Group A consists of isolates 01, 02, 08, 15, 16, 17, and 28; group B consists of isolates 22, 23, 39, 67, 76, 88, 104, and 105; and group C consists of isolates 73, 87, and 101. Typical colonies were characterized by large, gummy, and pale color with or without pink center. E. cloacae clinical isolate was used as a positive control.

Colony morphology of the Enterobacter isolates on MacConkey agar plates. Three morphological groups identified as Groups A, B, and C with circular, raised, entire, and pink center morphotypes are shown in
Antibiogram of poultry Enterobacter isolates
A total of 18 morphologically and biochemically characterized presumptive Enterobacter spp. were tested against 19 commonly used antimicrobials belonging to 15 different groups of antibiotics. All of the 18 isolates were resistant to penicillin and rifampicin (Fig. 2). Three morphological groups of Enterobacter spp. were distinctly different in their antibiotic resistance pattern (Table 1). Group A was mostly resistant among three groups. Most of the isolates of this group were resistant to eight antibiotics: ampicillin, clindamycin, erythromycin, nitrofurantoin, imipenem, penicillin, streptomycin, and vancomycin, whereas Group B isolates showed resistance to six antibiotics except two: streptomycin and moderately resistant to nitrofurantoin. In contrast, in addition to six antibiotics that were shown resistant by Group A and B isolates, Group C were resistant to sulfamethoxazole and sensitive to imipenem and streptomycin. However, most of them were resistant to 6–11 groups of antibiotics (Table 1).
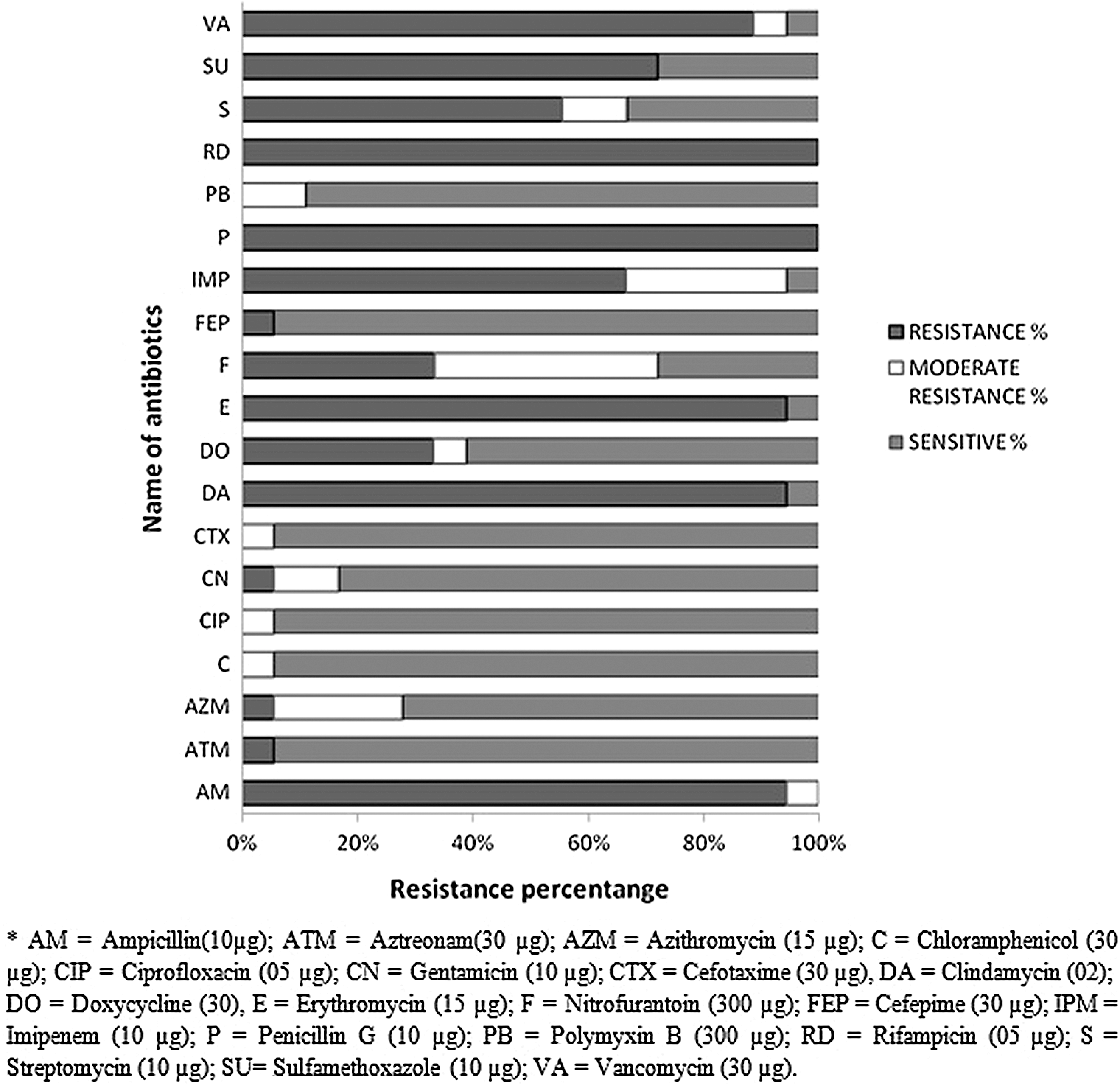
Antibiotic resistance pattern of the isolated Enterobacter spp. from poultry samples. The antibiotics used in this study are represented vertically and corresponding percentages of resistance, moderate resistance, and sensitive pattern of the Enterobacter isolates are represented horizontally.
AM, ampicillin (10 μg); ATM, aztreonam (30 μg); AZM, azithromycin (15 μg); C, chloramphenicol (30 μg); CIP, ciprofloxacin (5 μg); CN, gentamicin (10 μg); CTX, cefotaxime (30 μg); DA, clindamycin (2 μg); DO, doxycycline (30 μg); E, erythromycin (15 μg); F, nitrofurantoin (300 μg); FEP, cefepime (30 μg); IPM, imipenem (10 μg); P, penicillin G (10 μg); PB, polymyxin B (300 μg); RD, rifampicin (05 μg); S, streptomycin (10 μg); SU, sulfamethoxazole (10 μg); VA, vancomycin (30 μg).
Analysis of antibiotic resistance gene
Isolates were selected for specific antibiotic resistance gene PCR according to the antibiogram profile determined before (Table 1). None of the selected isolates showed amplification product of quinolone resistance or extended-spectrum beta-lactamase (ESBL)-specific genes. All of the isolates except 87 showed characteristic 377-bp amplification product of vancomycin resistant gene and 13 isolates showed characteristic 169-bp amplification product of sulfonamide resistant gene.
Molecular characterization and genotyping of the Enterobacter spp.
Plasmid profile analysis of the isolates revealed that all of the isolates were plasmid free because both of the methods used in the present investigation successfully extracted plasmid from E. coli V517.
ARDRA of a PCR product of approximately 900 bp of the Enterobacter spp. isolates using restriction enzyme HaeIII showed the same restriction pattern for all of the 18 Enterobacter isolates (Supplementary Fig. S1). Three fragments were obtained: 200 bp, 300 bp, and 400 bp. The RAPD analysis was carried out with two random primers, 1283 and R2 sequences in a single reaction. Both primers were able to discriminate different amplification patterns among the 18 isolates and deduced three different patterns (Fig. 3). Results with both primers correlated well with respect to differentiation of three individual genotypes (Types 1, 2, and 3, in Fig. 3). With 1283 and R2 primers, genotype 1 yields similar pattern for isolates 1, 2, 5, 15, 16, 17, 28; genotype 2 includes isolates 22, 23, 39, 67, 104, 105; and genotype 3 contains isolate 73, 87, 101 (Fig. 3). The result was also congruent with the three morphological groups, group A, B, C detected earlier.
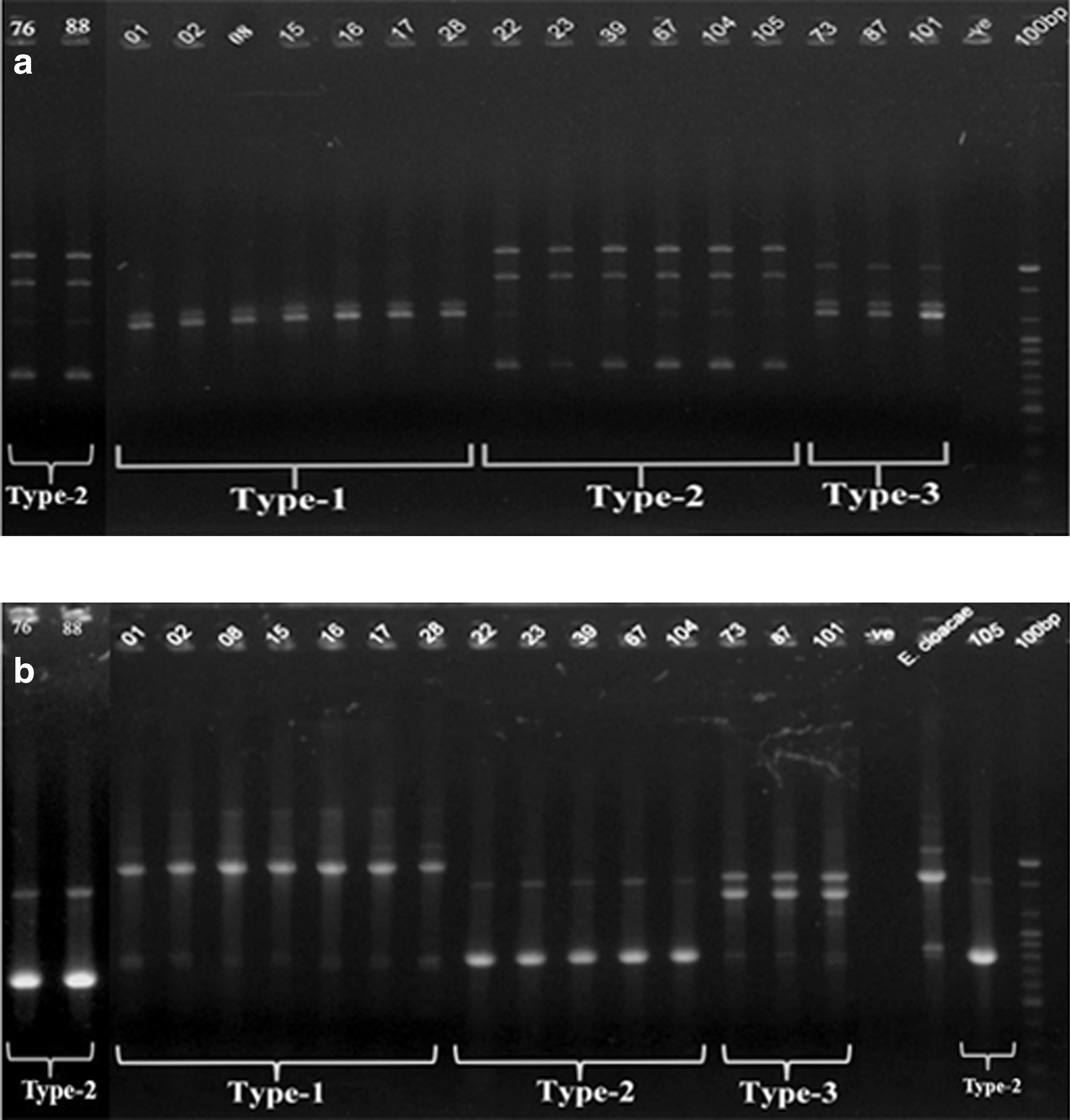
Randomly amplified polymorphic DNA genotyping profile by using primers 1283
Sequencing, phylogenetic analysis, and zoonotic correlation of the Enterobacter spp.
From each of the three genotypes, representative isolates were randomly selected for detailed 16S rRNA gene sequence analysis. From genotype 1, 2, and 3, isolates selected were 15 and 28; 39 and 104; and 101, respectively.
A phylogenetic tree was constructed in MEGA5 software using Neighbour-Joining algorithm and 1000 bootstrap replicates to deduce close phylogenetic relationship based on 16S rRNA gene sequences analysis of the three genotypic Enterobacter spp. (Fig. 4). Isolate 15 from genotype 1 was phylogenetically closely related to E. hormaechei strainLRC5 (JF772054) with a similarity of 100%. Isolate 28 from genotype 1 was phylogenetically closely related to E. hormaechei strain WW2 (JN993998) with a similarity of 99%. Isolate 39 from genotype 2 was phylogenetically closely related to Enterobacter cloacae strain SJ6 (EU779827) with a similarity of 99%. Isolate 104 from genotype 2 was phylogenetically closely related to E. cloacae subsp. dissolvens (NR044987) with a similarity of 100%. Finally, isolate 101 from genotype 3 was phylogenetically closely related to E. cancerogenus strain NB141A (JN644583) with a similarity of 99% phylogenetically; poultry Enterobacter spp. were distributed within two distinct clusters (Fig. 4). Genotypes 1 and 3 within the E. hormaechei/cancerogenus cluster and genotype 2 within the E. cloacae cluster.

Phylogenetic tree of 16S rRNA gene sequences of Enterobacter spp. isolates and close relative reference isolates retrieved from database with accession numbers. Sequences of the isolates obtained in this study are shown in bold, underlined. Corresponding randomly amplified polymorphic DNA types are mentioned for the isolates. The tree was generated in program MEGA 5 using the Neighbour-Joining algorithm, with the Spirochete sequences serving as outgroup. Bootstrap values (n=1000 replicates) are shown at branch nodes and the scale bar represents the number of changes per nucleotide position.
Discussion
Enterobacter spp. are important opportunistic pathogens that can cause nosocomial outbreaks and invasive infections such as bloodstream infections (Hidron et al., 2008). The prevalence of MDR Enterobacter spp. carrying ESBLs and plasmid-mediated quinolone resistance has been increasing worldwide (Kanamori et al., 2012). The present study found an overall 17% prevalence of three morphotypes (Fig. 1) of Enterobacter spp. within the poultry isolates of five different farms at Savar areas, probably due to their intrafarm transmission. All the Enterobacter isolates were found to be resistant to rifampicin and penicillin-G (Fig. 2), followed by ampicillin, erythromycin, clindamycin, vancomycin, sulfamethoxazole, and imipenem. Streptomycin and doxycyclines were moderate resistance. Chloramphenicol, cefepime, cefotaxime, ciprofloxacin, and gentamicin were sensitive to most of the isolates tested. The MDR profile of Enterobacter isolates might be due to overuse and misuse of antibiotics in poultry and indicates a potential threat to public health.
A similar antibiotic resistance profile was observed for E. cloacae isolated from environmental samples in Europe, where all of the E. cloacae studied were resistant to ampicillin, erythromycin, rifampicin, and sulfamethoxazole (Cornaglia et al., 2004). Also, in 2009, all of the E. cloacae, collected at Charles Nicolle hospital in Tunisia, showed resistance to fluoroquinolones, gentamicin, tobramycin, and trimethoprim-sulfamethoxazole and were variably resistant to netilmicin, amikacin, and tetracyclines (Hammami et al., 2012). In another study, various degrees of resistance of enterobacteria (including Enterobacter spp.) were observed, but they were susceptible to chloramphenicol and gentamicin (Haryani et al., 2008). In our study, all of the poultry Enterobacter spp. were sensitive to chloramphenicol, ciprofloxacin, and gentamicin. It has been reported that half of the world's antibiotic production is used in farm animals, resulting in development of resistant bacteria (Phillips et al., 2004). The MDR Enterobacter isolates from poultry showed amplification products, with the vanA gene responsible for vancomycin resistance and sulI genes responsible for sulfonamide resistance. None of the other genes tested were found to be positive. Perhaps PCR with other specific antibiotic resistance gene primers is required to determine the pool of antibiotic resistance gene within the Enterobacter isolates.
On the other hand, all of the MDR Enterobacter isolates were found to be plasmid free using both kit and manual plasmid extraction methods. The experimental procedures were within the limit of acceptance because plasmid was successfully extracted from the positive control E. coli V517. Plasmid-free E. cloacae were previously reported from street foods in Malaysia (Haryani et al., 2008). Plasmid-free MDR Salmonella were reported previously from Bangladesh poultry samples (Mahmud et al., 2011). The plasmid-free property of the poultry MDR-Enterobacter isolates may indicate that the genes encoding resistance to these antibiotics are chromosomal borne or there may be other significant mechanisms behind these resistances that are yet to revealed.
The present study employed DNA fingerprinting methods such as ARDRA and RAPD to detect genotypic variation within the poultry Enterobacter spp. and finally the 16S rRNA gene sequencing approach to characterize the isolated Enterobacter spp. ARDRA with HaeIII enzyme of the approximate 900-bp amplified fragment of 16S rRNA gene showed an identical restriction pattern of the analyzed isolates with varying morphological types. ARDRA was done only with one restriction enzyme of HaeIII. ARDRA with HaeIII enzyme showing single restriction pattern could be used as a reliable method to identify Enterobacter spp. at the genus level, but it may not be useful for intraspecies differentiation.
Another genotypic method, RAPD, enabled the elucidation of the genetic diversity among the isolates and within Enterobacter species. Three different RAPD types were obtained at a similarity level of 100%, for which RAPD PCR was useful not only for strain typing, but also for species identification of the isolates. The three RAPD types—Types 1, 2, and 3—also correlated well with the three morphological types—Groups A, B, and C, respectively—on MacConkey agar plates. Therefore, a combination of morphological, plasmid pattern, ARDRA, and RAPD approaches successfully differentiated the presence of three phenotypic as well as genotypic Enterobacter spp. prevalent in the poultry of Bangladesh.
The most prevalent species observed within the poultry of Bangladesh through phenotypic and genotypic characterization were E. cloacae, E. hormaechei, and E. cancerogenus. By 16S rRNA gene sequencing, it was determined that RAPD Type 2 was phylogenetically related to E. cloacae, while Type 1 and Type 3 have divergent genomes that more closely resembled that of E. hormaechei and E. cancerogenus, respectively. These species are commonly reported from clinical samples (Morand et al., 2009; Paauw et al., 2009), patients with cancer (Garazzino et al., 2005), animals (Sidjabat et al., 2007), and wholesome broiler carcasses (Moreira et al., 2009). Therefore, isolation and presence of such strains in poultry samples are indicative of zoonotic hazard.
From the data, it is evident that various Enterobacter spp. are emerging as increasingly important pathogens that possess innate resistance to older antimicrobial agents and are rapidly developing resistance to newer agents (Sanders and Sanders, 1997). The emergence of MDR zoonotic Enterobacter spp. in poultry of Bangladesh depicts the alarming situation of overuse or misuse of antibiotics in the poultry feed and indicates the necessity of good management practices in poultry.
Conclusions
The present study is a first endeavor to detect the prevalence and emergence of MDR Enterobacter spp. bearing clinical significance of zoonoses from poultry samples of five selected poultry farms at Savar in Bangladesh. Three morphotypes of Enterobacter spp. are prevalent (17%) within the poultry of five different farms at Savar, Dhaka. All of the poultry Enterobacter isolates are MDR at alarming levels, showing resistance to penicillin, rifampicin, erythromycin, clindamycin, sulfamethoxazole, and even imipenem. The MDR properties of Enterobacter isolates may have chromosomal origin. The prevalence of E. cloacae, E. hormaechei, and E. cancerogenus, which are phylogenetically close to the potentially pathogenic Enterobacter spp., indicates that zoonotic transmission of these strains could be a potential threat to public health.
Footnotes
Acknowledgments
The authors would like to thank Professor S. I. Khan of the Department of Microbiology, University of Dhaka for providing Enterobacter cloacae clinical strain and RAPD primers. This study was done with a grant provided by the Ministry of Education and Ministry of Science and Information and Communication Technology, Government of Bangladesh.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.