
Abstract
This study analyzed various organic fertilizers for indicator microorganisms, pathogens, and antibiotic-resistant Escherichia coli, and evaluated the growth potential of E. coli O157:H7 and Salmonella in fertilizers. A microbiological survey was conducted on 103 organic fertilizers from across the United States. Moisture content ranged from approximately 1% to 86.4%, and the average pH was 7.77. The total aerobic mesophiles ranged from approximately 3 to 9 log colony-forming units (CFU)/g. Enterobacteriaceae populations were in the range of <1 to approximately 7 log CFU/g, while coliform levels varied from <1 to approximately 6 log CFU/g. Thirty samples (29%) were positive for E. coli, with levels reaching approximately 6 log CFU/g. There were no confirmed positives for E. coli O157:H7, Salmonella, or Listeria monocytogenes. The majority of E. coli isolates (n=73), confirmed by glutamate decarboxylase (gad) PCR, were from group B1 (48%) and group A (32%). Resistance to 16 antibiotics was examined for 73 E. coli isolates, with 11 isolates having resistance to at least one antibiotic, 5 isolates to ≥2 antibiotics, and 2 isolates to ≥10 antibiotics. In the presence of high levels of background aerobic mesophiles, Salmonella and E. coli O157:H7 grew approximately 1 log CFU/g within 1 day of incubation in plant-based compost and fish emulsion-based compost, respectively. With low levels of background aerobic mesophiles, Salmonella grew approximately 2.6, 3.0, 3.0, and 3.2 log CFU/g in blood, bone, and feather meals and the mixed-source fertilizer, respectively, whereas E. coli O157:H7 grew approximately 4.6, 4.0, 4.0, and 4.8 log CFU/g, respectively. Our results revealed that the microbiological quality of organic fertilizers varies greatly, with some fertilizers containing antibiotic resistant E. coli and a few supporting the growth of foodborne pathogens after reintroduction into the fertilizer.
Introduction
O
There is also concern worldwide about the increased prevalence of antimicrobial resistance in microbial communities. The use of antimicrobials in food animal production to prevent disease and promote growth is considered the main risk factor for increased resistance in pathogenic bacteria (van den Bogaard and Stobberingh, 2000; McEwen and Fedorka-Cray, 2002). These resistant bacteria can be introduced into a farm environment through animal feces (Carson et al., 2008; Jiang et al., 2006; Smith et al., 2007; Yang et al., 2006) and spread to agricultural crops (Campagnolo et al., 2002). Antimicrobial resistance genes can be transferred horizontally to other strains or species in the gut of livestock as well as in manure and composted animal wastes (Götz and Smalla, 1997; Blake et al., 2003; Guan et al., 2007). Most importantly, antimicrobial resistance genes can be transferred from commensal bacteria to pathogenic strains (Blake et al., 2003).
There are few reports on the microbiological quality of organic fertilizers. Brinton et al. (2009) surveyed 94 market-ready recycled organic composts in Washington, Oregon, and California and found that 23%, 44%, and 20% of samples exceeded the Environmental Protection Agency (EPA) 503 limit of 1000 most probable number (MPN)/g for fecal coliforms, respectively (US NRAR, 1997). One sample from Washington was positive for Salmonella, and samples from three composting facilities in California were positive for E. coli O157:H7. Lasaridi et al. (2006) reported that most of compost samples (n=28) from the Greek market had background populations>108 colony-forming units (CFU)/g, whereas fecal coliforms exceeded 103 CFU/g in all samples. No Salmonella was found in any of the samples tested.
Thermophilic composting is used to reduce the occurrence of pathogens from animal waste products. However, the composting process may not reduce pathogens to acceptable levels, or pathogens may be reintroduced into the finished product by cross-contamination on the farm. Studies show that Salmonella and E. coli grew and survived longer in compost with the moisture content above 20% (Hussong et al., 1985; Pietronave et al., 2004). E. coli, Salmonella, E. coli O157:H7, and Listeria monocytogenes all had greater growth potential in sterilized compost as compared to nonsterilized compost (Yeager and Ward, 1981; Sidhu et al., 2001; Kim and Jiang, 2010).
This study aimed to analyze organic fertilizer samples collected from multiple states microbiologically, determine the presence of antimicrobial resistance in E. coli isolates, and evaluate growth potential of both E. coli O157:H7 and Salmonella in selected organic fertilizers.
Materials and Methods
Sample collection
Samples of organic fertilizers were collected aseptically from multiple states in the United States including Arizona, California, Georgia, Kentucky, Maryland, New York, North Carolina, South Carolina, and Tennessee from 2007 to 2010. The main components—age of compost and location collected—were recorded. Samples were gathered from nearby farms, some were shipped from outside locations, and others were bought commercially. The samples were transported at ambient temperatures and stored in a refrigerator (5°C) upon arrival.
Water activity, moisture content, and pH measurements
The water activity of each sample was measured using a dew-point water activity meter (Aqualab series 3TE, Decagon Devices, Pullman, WA). Moisture content and pH were determined as described previously (Shepherd et al., 2007).
Microbiological analysis
Twenty-five grams of each organic fertilizer were added to 225 ml of universal pre-enrichment broth (UPB; Neogen, Lansing, MI) in a Whirl-Pak® sampling bag and shaken vigorously. For enumeration of background aerobic mesophiles, serial dilutions were made from each UPB suspension, spiral-plated on tryptic soy agar (Becton & Dickinson, Sparks, MD), and incubated overnight at 37°C. Enterobacteriaceae was enumerated in violet red bile agar with glucose (Becton & Dickinson) by pour-plating, then overlaid with violet red bile agar and incubated overnight at 37°C. Coliforms and E. coli were enumerated by plating 1 mL of UPB suspension onto E. coli/coliform Petrifilm® (3M, St. Paul, MN), and incubated overnight at 37°C. The remaining UPB suspensions were incubated overnight at 37°C with shaking for pathogen detection as described below.
Pathogen detection
Each overnight culture of UPB was transferred to Rappaport-Vassiliadis broth (Becton & Dickinson), modified tryptic soy broth (Becton & Dickinson) with novobiocin (20 μg/mL) (Oxoid Ltd., Basingstoke, Hants, UK), and Fraser broth (Becton & Dickinson) for selective enrichment of Salmonella, E. coli O157:H7, and L. monocytogenes, respectively, and incubated overnight at 37°C with shaking. Samples were then streaked onto sorbitol MacConkey agar (Becton & Dickinson), Oxford agar (Becton & Dickinson), and xylose-lysine-tergitol 4 agar (Becton & Dickinson) for detection of E. coli O157:H7, L. monocytogenes, and Salmonella, respectively. Presumptive positive colonies of the target pathogen were confirmed by real-time polymerase chain reaction (PCR) as described previously by Heringa et al. (2010).
Characterization of E. coli isolates
Three to five typical E. coli colonies (blue with gas bubble formation) per sample were picked from all positive Petrifilms® and streaked on tryptic soy agar until pure colonies were obtained. The DNA of presumptive E. coli isolates was extracted using boiling method. The primers listed in Table 1 were synthesized by Invitrogen Co. (Carlsbad, CA). Presumptive E. coli isolates were verified by real-time PCR of the glutamate decarboxylase gene (gad) as previously described by Chen et al. (2006). The SYBR green-based method was performed using the Bio-Rad iCycler™ system (BioRad, Inc., Hercules, CA). A triplex PCR was carried out for phylogenetic analysis of E. coli isolates as described by Clermont et al. (2000) with some modifications (Heringa et al., 2010).
gad, glutamate decarboxylase; stx1 and 2, Shiga toxins 1 and 2; chuA, yjaA, and TSPE4.C2, phylogenetic group markers; rfbE, enzyme necessary for O-antigen biosynthesis; hly, hemolysin; fimI, fimbrin-like protein.
Antimicrobial susceptibility testing
All E. coli isolates were tested for susceptibility to a series of antimicrobial agents on Mueller Hinton agar (Difco) using agar dilution method according to procedures described by the Clinical and Laboratory Standards Institute (CLSI, 2009). The antimicrobial agents (n=16) tested were chosen based on the National Antimicrobial Resistance Monitoring System including amikacin, ampicillin, amoxicillin/clavulanic acid, ceftriaxone, chloramphenicol, ciprofloxacin, trimethoprim/sulfamethoxazole, cefoxitin, gentamicin, kanamycin, nalidixic acid, sulfisoxazole, streptomycin, tetracycline, ceftiofur, and cefotaxime (Sigma). The minimum inhibitory concentration results were interpreted by the use of CLSI breakpoints for Enterobacteriaceae. E. coli 25922 (ATCC, Manassas, VA) was used as the quality control strain for antimicrobial susceptibility testing. Each experiment was performed in triplicate.
Bacterial cultures
A three-strain mixture of both E. coli O157:H7 and Salmonella was used as the inocula for regrowth study. Strains F06M-0923-21 (Spinach outbreak, from California Department of Health), F07M-020-1 (Taco John outbreak, from California Department of Health), and B6914 (stx 1- and stx 2-, from Dr. Fratamico, USDA-ARS-ERRC) were selected for E. coli O157:H7, as well as Salmonella serovars Enteritidis H2292 (from Dr. Mike Doyle, University of Georgia), Heidelberg 21380 (from Dr. Shaohua Zhao, CVM-FDA), and avirulent Typhimurium 8243 (from Dr. Roy Curtis, Washington University). These strains were induced to be resistant to 100 μg rifampin per milliliter (Fisher Scientific, Fair Lawn, NJ) through the gradient plate method (Rice and Bonomo, 2005) and stored at −80°C in tryptic soy broth with 20% glycerol.
Growth potential of pathogens in organic fertilizers
The growth potential of E. coli O157:H7 and Salmonella was evaluated in fertilizers with two levels of background aerobic mesophiles. The organic fertilizers with a high level of aerobic mesophiles (≥5 log CFU/g) tested for potential growth of E. coli O157:H7 included fish emulsion–based compost (n=1), cow manure–based compost (n=2), and spent mushroom compost (n=2). The growth potential of Salmonella was evaluated separately in each of the following fertilizers (n=5): spent mushroom compost, super-heated chicken litter, plant-based compost, horse manure–based compost, and hen manure–based compost. Moisture content was adjusted to 40% for the composted fish emulsion and 50% for super-heated chicken litter in order to achieve a water activity above 0.95. All other organic fertilizers were used at their original moisture content.
The same organic fertilizers with a low level of background aerobic mesophiles (<5 log CFU/g) were used for both pathogens including bone meal, blood meal, feather meal, cow manure–based compost (pH=10), and a mixed-source fertilizer, whereas a cow manure–based compost (pH=7) with a high level of aerobic mesophiles (approximately 7 log CFU/g) was used as a control. All samples were adjusted to moisture contents between 30% and 50% prior to pathogen inoculation to yield water activities of at least 0.95.
An inoculum cocktail of three rifampin-resistant strains of E. coli O157:H7 or Salmonella was sprayed onto compost to a final concentration of approximately 5 log CFU/g using a sterile spray nozzle. The compost was then mixed by hand while wearing sterile gloves until the inoculum was evenly distributed. The samples were stored in sampling bags at 22°C or 30°C, and taken at selected intervals to determine pathogen populations. Each growth study was conducted for two trials.
Statistical analysis
Bacterial counts were converted to log CFU/g dry weight and subjected to analysis of variance using SAS (ver. 9.1; SAS Institute, Inc., Cary, NC). P-value below 0.05 was considered significantly different. Tukey's honestly significantly different test was used to determine whether the growth of pathogens differed at each sampling time.
Results
Analysis of organic fertilizers
A total of 103 organic fertilizer samples were collected from multiple states across the United States (Table 2). The moisture content ranged from approximately 1% to 86.4% with water activities between 0.298 and 0.999. The vermicompost group had the highest average moisture content at approximately 75%, while the bone, blood, and feather meals had the lowest average at 8.1%. The spent-mushroom composts, vermicomposts, alpaca manure, and mixed animal waste composts all had an average water activity above 0.99. The pH of most samples was neutral with the average pH at 7.8. However, the pH of liquid fish emulsion samples (n=4) was around 3–4, as compared with pH of approximately 10 for cow manure–based composts (n=2) collected from Arizona.
Percent positive among samples tested in each category.
NA: not enough data to calculate a median.
SD, standard deviation; CFU, colony-forming units; TBC, total bacterial count.
The counts of aerobic mesophiles ranged from approximately 3.0 to 9.0 log CFU/g with the highest average counts of approximately 8.0 log CFU/g from horse manure–based compost and rabbit manure–based compost groups. Both liquid fish emulsion and blood, bone, and feather meal groups had lower average aerobic mesophile counts at approximately 4.0 log CFU/g. Enterobacteriaceae populations were in the range of <1 to 7.0 logs CFU/g. All vermicompost (n=5), rabbit manure–based compost (n=3), greenwaste (n=9), and alpaca manure samples (n=2) were positive for Enterobacteriaceae. Coliform levels varied from <1.0 to approximately 6.0 logs CFU/g with an average of 3.4 logs CFU/g. Thirty samples (29%) were found to be positive for E. coli, with the largest proportion of those coming from horse manure–based composts. The average number of E. coli ranged from 2.7 to 6.3 log CFU/g. The super-heated chicken litter, liquid fish emulsion, composted fish emulsion, and plant-based compost were all negative for Enterobacteriaceae, coliforms, and E. coli. Twenty-seven samples were considered as presumptive positive for Listeria spp. However, no samples were confirmed positive for E. coli O157:H7, Salmonella, or L. monocytogenes after species-specific PCR analysis.
E. coli characterization
Eighty-eight putative E. coli colonies were picked and 73 were confirmed as E. coli by gad PCR (Table 3). PCR was used further to determine the phylogenetic groups of E. coli isolates with a 94% positive identification rate. The majority of isolates belonged to group B1 (n=35, 48%) and group A (n=23, 32%), compared with groups B2 (n=3, 4%) and D (n=7, 9%). The isolates were also screened for Shiga toxins 1 and 2 by real-time PCR, but none was positive for either virulence gene.
Confirmed by both Gram staining and glutamate decarboxylase–polymerase chain reaction.
AMP, ampicillin; AUG, amoxicillin/clavulanic acid; AXO, ceftriaxone; CHL, chloramphenicol; SXT, trimethoprim/sulfamethoxazole; FOX, cefoxitin; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; STR, streptomycin; TET, tetracycline; TIO, ceftiofur; CTX, cefotaxime. The resistance breakpoints for the tested antibiotics were 4 μg/mL for CTX; 8 μg/mL for TIO; 16 μg/mL for GEN and TET; 32 μg/mL for AMP, CHL, FOX, and NAL; 64 μg/mL for AXO, KAN, and STR; 4/76 μg/mL for SXT; and 32/16 for AUG.
Antibiotic susceptibility of E. coli isolates
Resistance to 16 antibiotics was examined for all 73 confirmed E. coli isolates (Table 3). Eleven isolates were resistant to at least one antibiotic, five isolates were resistant to ≥2 antibiotics, and two isolates were resistant to ≥10 antibiotics. The largest percentage of resistant isolates was from poultry manure–based compost (27%), whereas no isolates from the mixed animal compost or the mixed source fertilizer had resistance. Two isolates that showed resistance to more than 10 antibiotics were from the alpaca manure and the poultry manure–based compost samples. The majority of isolates with antibiotic resistance were resistant to cefoxitin (45%), ampicillin, tetracycline, and streptomycin (36% each). No isolates were resistant to amikacin, ciprofloxacin, or sulfisoxazole.
Growth potential of E. coli O157:H7 and Salmonella in various organic fertilizers with high levels of background aerobic mesophiles
E. coli O157:H7 was tested for growth potential in organic fertilizers with ≥5 log CFU/g of aerobic mesophiles at both 22°C and 30°C for 7 days (Fig. 1A and B and Table 4). E. coli O157:H7 had significant growth (p<0.05) of approximately 1 log within 1 day at both 22°C and 30°C in the composted fish emulsion sample only. Since growth potential for E. coli O157:H7 was not different at 22°C than 30°C, the following growth potential studies were conducted at 22°C only.
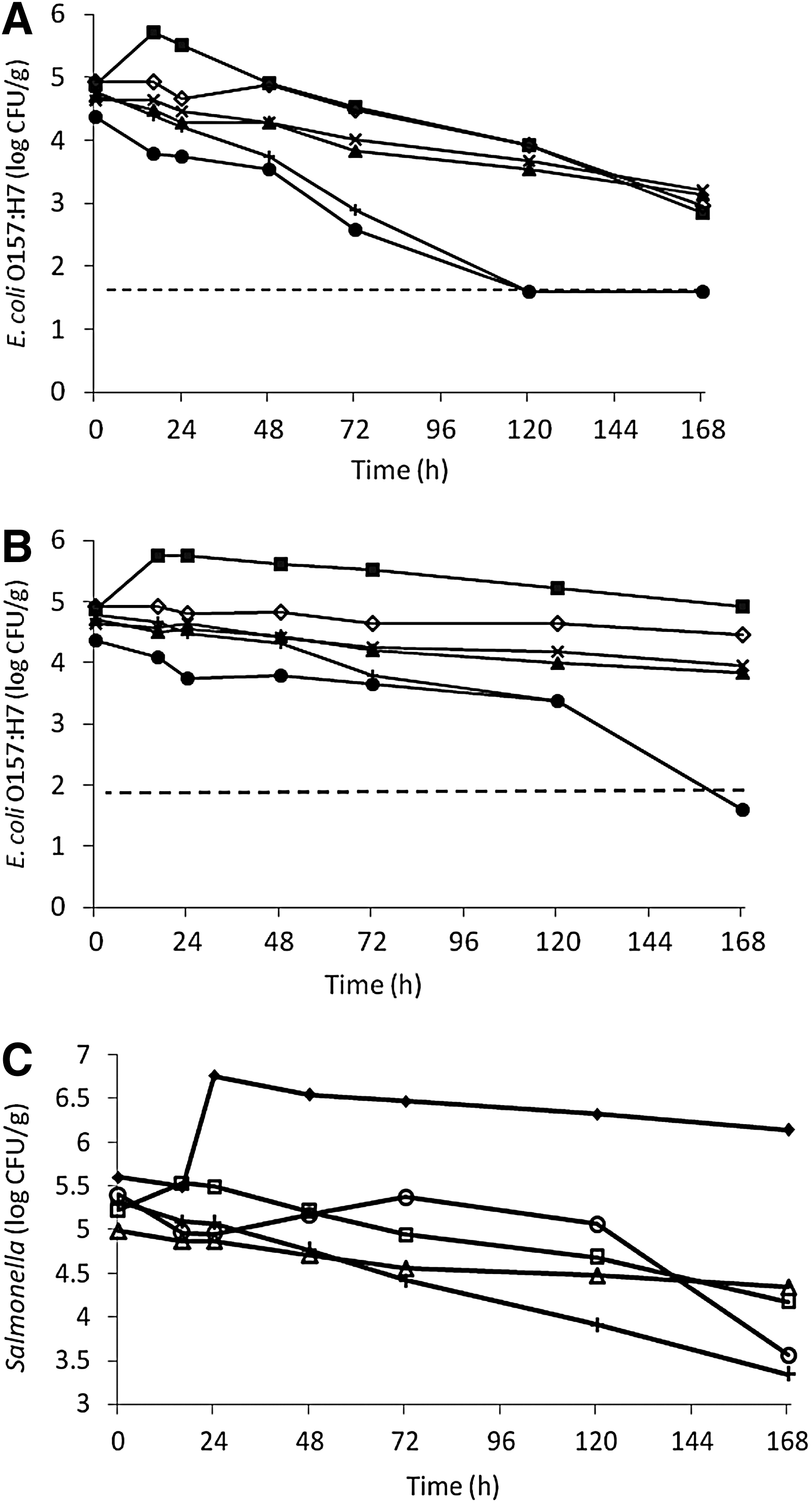
Growth potential of Escherichia coli (E. coli) O157:H7 at 30°C
Growth potential was defined as the difference of plate counts from the inoculum level.
SD, standard deviation; CFU, colony-forming units; +, growth; —, no growth; N/A, not tested.
Growth potential of Salmonella in five different types of organic fertilizers with ≥5 log CFU/g of background aerobic mesophiles was tested at 22°C (Fig. 1C and Table 4). At initial inoculum level of approximately 5 log CFU/g, Salmonella grew (p<0.05) by approximately 1 log within 1 day in plant-based compost and then stayed at a steady state until day 7.
Growth potential of E. coli O157:H7 and Salmonella in various organic fertilizers with low levels of background aerobic mesophiles
Figure 2A and B presents data on growth potential of E. coli O157:H7 and Salmonella in organic fertilizers with <5 log CFU/g of aerobic mesophiles at 22°C (Table 4). At initial inoculum levels of approximately 5 logs CFU/g, E. coli O157:H7 and Salmonella grew significantly (p<0.05) in all samples except for two cow manure–based composts. Within 7 days of incubation, E. coli O157:H7 populations increased approximately 4.6, 4.0, 4.8, and 4.0 log CFU/g in bone meal, blood meal, mixed source fertilizer, and feather meal, respectively, while Salmonella populations increased approximately 2.6, 3.0, 3.2, and 3.0 logs CFU/g, respectively.
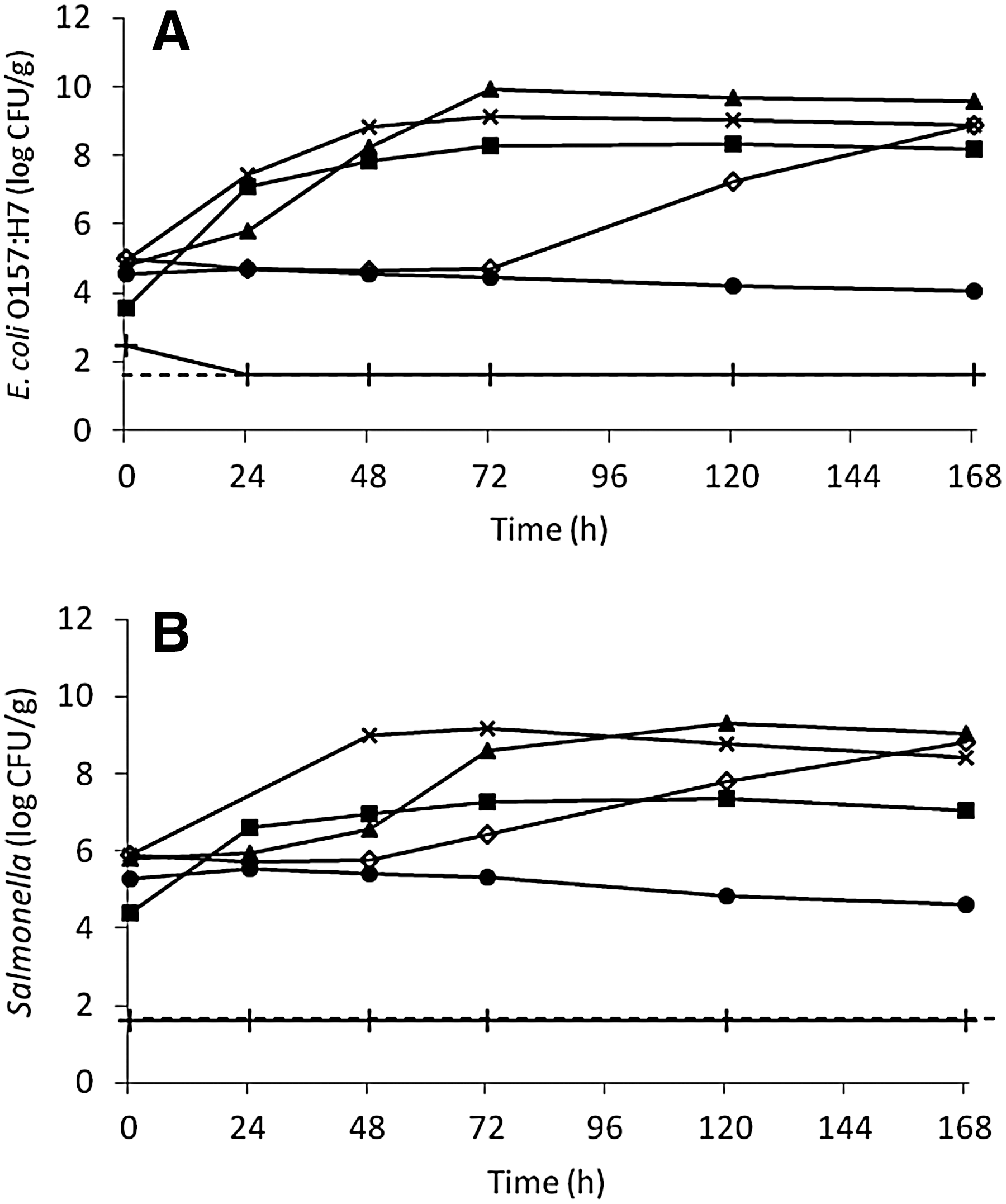
Growth potential of Escherichia coli (E. coli) O157:H7
Discussion
Most organic fertilizers analyzed in this study went through some types of heat treatment such as thermophilic composting, dry-heat for super-heated chicken litter, and boiling for fish emulsion. Therefore, the finished organic fertilizers should have very low levels of potentially pathogenic bacteria such as E. coli and other indicator microorganisms. Our survey found 30 out of 103 organic fertilizer samples positive for E. coli (29%). However, none of organic fertilizer samples analyzed in this study were positive for Salmonella, E. coli O157:H7, and L. monocytogenes. Brinton et al. (2009) surveyed 94 compost samples, with one third of the samples having E. coli levels less than 10 MPN/g and another one third exceeding 1000 MPN/g. One out of 94 samples was positive for Salmonella but was below the Environmental Protection Agency limit of 3 MPN/4 g. E. coli O157:H7 was detected in samples from three large composting facilities. The above results indicate that if optimal conditions are not met, the composting process may not be adequate to eliminate all pathogenic bacteria.
Fecal coliforms such as E. coli are useful indicators of compost hygiene (Brinton et al., 2009; Elving et al., 2009), and antimicrobial profiles for E. coli may give some insight into the development of resistance in other foodborne pathogens. Also, the abundance of antimicrobial-resistant E. coli implicates them as likely vehicles for the spread of resistance genes (O'Brien, 2002).
Our results on antimicrobial resistance in E. coli seem to agree with published data that resistance to older antibiotics is more widespread than resistance to newer drugs (Prescott et al., 2000). For example, there were higher numbers of resistant isolates to older aminoglycosides than to newer antibiotics, with streptomycin, kanamycin, gentamicin, and amikacin having 4, 2, 1, and 0 resistant isolates, respectively. Antimicrobial resistance also seems to be more common for antibiotics frequently used to treat farm animals. In this study, we found four isolates each with resistance to ampicillin and tetracycline, which are broad-spectrum antibiotics commonly used to treat animals (Prescott et al., 2000; Wilson, 2001). Dairy/cattle farms use many first- and second-generation cephalosporins, while the only third-generation cephalosporins approved for use are cefoperazone and ceftiofur. In this study, five isolates had resistance to cefoxitin, a second-generation cephalosporin, while ceftriaxone, cefotaxime, and ceftiofur, third-generation cephalosporins, had fewer resistant isolates with 2, 2, and 0, respectively.
Pathogens may get introduced into organic fertilizers after the thermophilic phase of composting or heating treatment is over. Insects and wild animals, water runoff, and equipment used in fertilizer handling may contaminate organic fertilizers. Since organic fertilizers are rich in nutrients, reintroduced pathogens may multiply under certain conditions. In this study, E. coli O157:H7 and Salmonella grew in certain organic fertilizers with growth ranging from 1 to 5 logs. Numerous studies have shown that Salmonella, E. coli O157:H7, and L. monocytogenes have a greater growth potential in sterilized compost as compared to nonsterilized compost (Yeager and Ward, 1981; Hussong et al., 1985; Sidhu et al., 2001; Kim and Jiang, 2010). Composts with a lower level of background microflora may support the growth of pathogens because there is less competition for water and nutrients. However, the suppressive effect of indigenous microflora also depends on the diversity of microorganisms present (Millner et al., 1987; Pietronave et al., 2004). Although plant-based compost in this study had high aerobic mesophilic bacterial counts (9.1 logs CFU/g), it still allowed growth of Salmonella. Therefore, unique biotic and abiotic characteristics may explain the growth of Salmonella in plant-based compost plus the growth of E. coli O157:H7 in fish emulsion–based compost. Further study is needed on analyzing microbial composition in compost.
In this study, only one inoculum concentration (approximately 5 logs CFU/g) was used to test the worst-case scenario of pathogen recontamination. However, a very low level of pathogen contamination is expected for real-world situations, and the growth potential of a few cells may be different from what we observed in this study due to the competitive nature of microorganisms. Therefore, further study is needed.
Conclusions
Based on our results, the microbiological quality of organic fertilizers varies greatly, and each sample is unique based on its specific biotic and abiotic characteristics. Overall, organic fertilizers analyzed in this study were free from the tested foodborne pathogens; however, E. coli was present in nearly 30% of samples analyzed. About 16% of E. coli isolates harbored antimicrobial resistance to at least one antibiotic. These results provide some baseline data on the quality of these organic fertilizer products. Furthermore, the growth of E. coli O157:H7 and Salmonella is more likely to occur in organic fertilizers containing low levels of indigenous microorganisms. Therefore, the microbiological safety of organic fertilizers, not only just animal waste-based composts, needs to be evaluated.
Footnotes
Acknowledgment
This project was funded by a grant from the Center for Produce Safety at the University of California, Davis.
Disclosure Statement
No competing financial interests exist.