
Abstract
Salmonella enterica subsp. enterica serovar 4,[5],12:i:- is a monophasic variant of Salmonella Typhimurium. In this study, a total of 651 human and 107 food and environmental isolates of serovar 4,[5],12:i:- recovered from 2007 through 2011 in Switzerland were characterized by antibiotic resistance profiles and pulsed-field gel electrophoresis (PFGE). In addition, a selection of isolates belonging to the most frequent PFGE patterns was further subjected to multilocus variable-number tandem-repeat analysis (MLVA) and phage typing. Over the years 2007–2011, the reports of salmonellosis caused by Salmonella enterica serovar 4,[5],12:i:- significantly increased. A high prevalence of multidrug-resistant isolates, mainly showing an ampicillin–streptomycin–sulfonamide–tetracycline resistance pattern (ASSuT), was observed. In addition, four extended spectrum beta lactamase (ESBL) (CTX-M-55)–producing isolates were found. XbaI PFGE analysis of all isolates revealed over 150 different pulsotypes, and generally showed a considerable diversity within the monophasic isolates. Nevertheless, among these we identified seven dominant profiles, which encompassed 66% of all isolates tested. The PFGE type STYMXB.0131 dominated among human as well as food isolates. Multilocus variable-number tandem-repeat analysis profile 3-12-10-0-0211, which, in many cases, coincided with PFGE type STYMXB.0131 and phage type DT193 were the most prevalent types found for the isolates further characterized by these typing methods. Our data provide strong evidence for a spread of two specific Salmonella serovar 4,[5],12:i:- clones (PFGE pattern STYMXB.0131, resistance type ASSuT) and (PFGE pattern STYMXB.0131, resistance type SSuT). In contrast to the human isolates, the pork/poultry isolates expressed predominantly the SSuT resistance type.
Introduction
N
Since the year 2009, S. enterica subsp. enterica serovar 4,[5],12:i:- ranks among the three most frequent serovars found in Switzerland within the isolates submitted to the National Centre for Enteropathogenic Bacteria and Listeria for serotyping. It is known to be mainly a monophasic variant of Salmonella Typhimurium with antigenic and genotypic similarities (Echeita et al., 2001). As with other enteric salmonellae, infections due to serovar 4,[5],12:i:- commonly manifest themselves as self-limiting gastroenteritis, but severe invasive infections can also occur, which call for antimicrobial treatment. In those cases, fluoroquinolones, trimethoprim-sulfamethoxazole, ampicillin, or third-generation cephalosporins come into consideration (Hohmann, 2001). The emergence and spread of resistant bacteria and subsequent treatment failure is a significant and increasing global public health problem. A large proportion of the European serovar 4,[5],12:i:- strains is resistant to ampicillin, streptomycin, sulfonamides, and tetracycline, a resistance pattern (R-type) that is abbreviated by ASSuT (Hopkins et al., 2010). The sources of isolates causing human infections often turned out to be pigs and pork products (Hauser et al., 2010; Rodríguez et al., 2012).
The objective of this study was to characterize human, food, and environmental isolates of serovar 4,[5],12:i:- collected from 2007 through 2011 in order to gain data about the epidemiology, phenotypic and genetic diversity, and susceptibility to antimicrobials of this serovar circulating in Switzerland. Such data provide a scientific basis for law-enforcement actions by food safety authorities.
Materials and Methods
Salmonella isolates
From 2007 through 2011, 8256 cases of Salmonella infection were reported to the Swiss Federal Office of Public Health. From this time span, 758 isolates of serovar 4,[5],12:i:- (651 from humans and 107 from food or environment [water] samples) were selected for this study. Most of the 651 human isolates were derived from stool samples. The remaining isolates originated from urine (11), blood (5), or other sites (44). More male (53.7%) than female cases (46.3%) were recorded. The average age for infection with Salmonella serovar 4,[5],12:i:- was 21 years (men: 19 years, women: 22 years). Multiple isolates from the same patient or isolates from family members were excluded. Nonhuman isolates (N=107) originated from pork (N=71), poultry (N=22), the environment (N=8), and for 6 isolates the origin was unknown. All samples had been collected and submitted to the National Centre for Enteropathogenic Bacteria and Listeria by diagnostic laboratories, hospitals, or family doctors throughout Switzerland.
Antibiotic susceptibility testing
The isolates were tested for antimicrobial susceptibility by the disk diffusion method according to the Performance Standards for Antimicrobial Susceptibility Testing (CLSI, 2008). The panel of antibiotic disks (Becton, Dickinson and Company, MD) consisted of ampicillin, amoxicillin/clavulanic acid, cephalothin, cefotaxime (CTX), ciprofloxacin, gentamicin, tetracycline, streptomycin, chloramphenicol, kanamycin, nalidixic acid, sulfamethoxazole, and trimethoprim. The strains were classified as resistant, intermediate, or susceptible to each antibiotic agent according to the CLSI criteria. For calculations of percentages, intermediate susceptibility was considered as resistant. Isolates displaying resistance to three or more antimicrobial compounds of different classes were defined as multidrug resistant.
Isolates whose resistance patterns demonstrated a synergy effect between amoxicillin/clavulanic acid and CTX in the disk diffusion test were further tested using E-Test extended spectrum beta lactamase (ESBL) strips (BioMérieux, Marcy l'Etoile, France) containing CTX, cefepime, or ceftazidime alone and in combination with clavulanic acid according to the manufacturer's protocols (Brown et al., 2000). Confirmed ESBL producers were analyzed for bla
TEM, bla
SHV, and bla
CTX-M genes by polymerase chain reaction (PCR) and by sequencing the whole open reading frames of the responsible bla genes, using specific primer sets (Pitout et al., 1998; Woodford et al., 2006; Geser et al., 2012). Resulting amplicons were purified using the PCR Purification Kit (Qiagen, Courtaboeuf, France) according to the manufacturer's recommendations. Custom sequencing was performed at Microsynth (Balgach, Switzerland) and the nucleotide and protein sequences were analyzed with Codon Code Aligner V. 3.7.1.1. For database searches, the BLASTN program of National Center for Biotechnology Information (
PFGE typing
Pulsed-field gel electrophoresis (PFGE) was performed using the restriction endonuclease XbaI (Roche, Mannheim, Germany) by following the Centers for Disease Control and Prevention (CDC) PulseNet protocol (
Multilocus variable-number tandem-repeat analysis (MLVA) and phage typing
Moreover, from the seven most frequent PFGE patterns, 40 isolates representing the genetic and antimicrobial resistance diversity were selected for MLVA and phage typing. MLVA was based on the protocol by Lindstedt et al. (2004) using an ABI 310 DNA Sequencer (Applied Biosystems, Darmstadt, Germany) (Beutlich et al., 2011). For data evaluation, nomenclature according to Larsson et al. (2009) was used. Assignation of the phage type was done according to the Anderson typing scheme (Anderson et al., 1977).
Results and Discussion
While the overall number of human salmonellosis cases in Switzerland decreased from 2001 to 2011, the number of isolates with serovar 4,[5],12:i:- increased during the last 8 years. In 2007, serovar 4,[5],12:i:- accounted for 5.8% (95% confidence interval [CI], 4.3; 7.6) of annual human Salmonella isolates. For the following years, those figures were as follows: 8.2% (95% CI, 6.7; 9.9) in 2008, 18.3% (95% CI, 15.9; 21.0) in 2009, 15.9% (95% CI, 13.6; 18.4) in 2010, and 18.2% (95% CI, 15.9; 20.7) in 2011, which was a significant change in the prevalence (p<0.05).
A total of 327 isolates (43%) showed one to three resistances and 424 isolates (56%) displayed more than three resistances, whereas only seven isolates (1%) were fully susceptible against the antibiotics tested. During the years 2007–2011, the trend shifted toward the ASSuT resistance pattern (Fig. 1). The prevalence of ASSuT significantly increased from 35.4% (95% CI, 22.2; 50.5) to 53.7% (95% CI, 46.3; 60.9) among the human isolates (p<0.05). We found 64% of all serovar 4,[5],12:i:- isolates resistant to ampicillin, 89% to streptomycin, 97% to sulfamethoxazole, and 88% to tetracycline while at most 6% of all isolates were resistant to any of the remaining nine antibiotics.
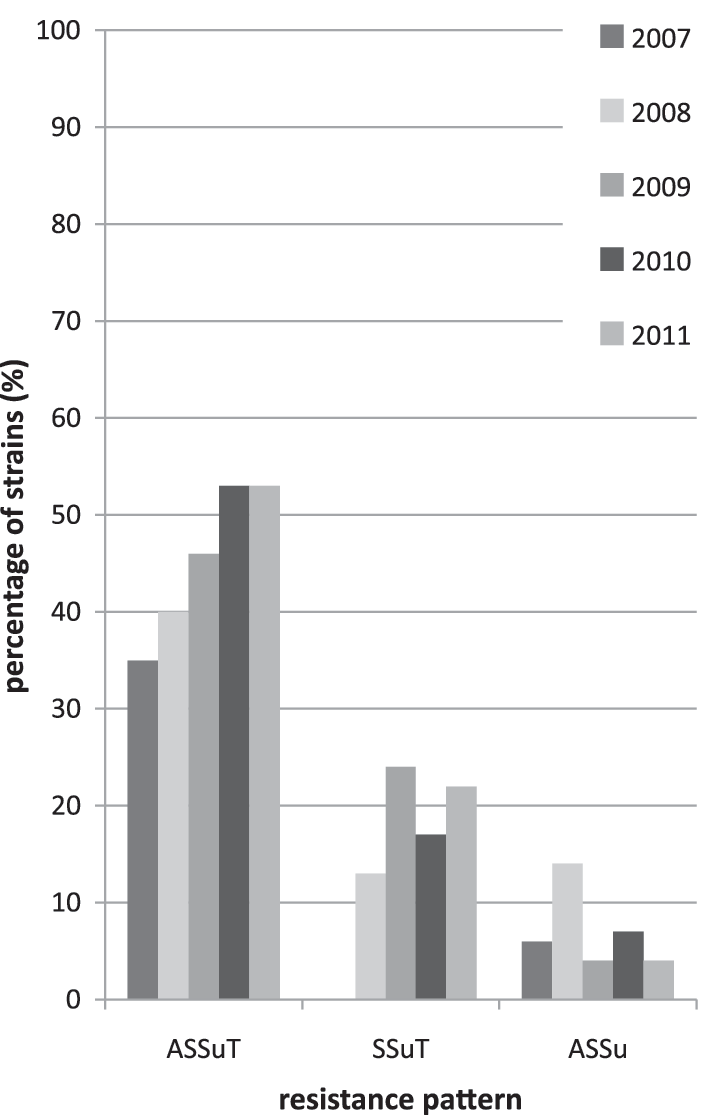
Percentage of human Salmonella enterica subsp. enterica serovar 4,[5],12:i:- isolates with the ampicillin–streptomycin–sulfonamide–tetracycline (ASSuT), streptomycin–sulfamethoxazole–tetracycline (SSuT), ampicillin–streptomycin–sulfonamide (ASSu) resistance patterns between 2007 and 2011.
The second most frequent pattern was resistance against SSuT, with none recorded in 2007 and 24% detected in 2009 among human isolates. However, the SSuT pattern was found most frequently among the nonhuman isolates (about 70%). The third most common pattern included resistance against ampicillin, sulfamethoxazole, and streptomycin (ASSu), and was found in about 7% of all human isolates tested, but never in nonhuman isolates.
Four human isolates expressed an ESBL phenotype (Table 1). The patients were 1, 22, and 23 years (two persons) of age. All four individuals lived in different cantons of Switzerland. These isolates were collected between 2009 and 2011. A comparison of their corresponding PFGE results showed different patterns for all four isolates (data not shown). They also differed from other serovar 4,[5],12:i:- XbaI PFGE patterns in Switzerland during this time period. PCR results revealed that all four isolates were positive for genes belonging to the TEM and the CTX-M families of β-lactamases. Sequencing of the entire open reading frames including flanking regions revealed the presence of bla TEM-1 and the ESBL gene bla CTX-M-55. Three of the four affected patients had a history of travel to Thailand shortly before disease onset. For one patient, there was no recorded history of foreign travel. Interestingly, CTX-M-55 was first reported from Thailand (Kiratisin et al., 2007), and—according to subsequent studies—remained endemic there. The fact that the four ESBL producers yielded various PFGE patterns despite the patients' similar anamnesis may well be explained by (1) the fact that ESBL genes in Salmonella are plasmid mediated and plasmids might spread independently to clonal spread of bacteria; (2) the long time period (3 years) during which they arose; and (3) the dynamics of the occurrence of blaCTX-M-55 in Thailand, a gene that was found there in Klebsiella pneumoniae, Escherichia coli, and also in Salmonella (Kiratisin et al., 2007), implying association with transferable plasmids.
XbaI PFGE analysis of the 758 isolates revealed over 150 different pulsotypes, and generally showed a considerable diversity within the monophasic isolates. Nevertheless, among these we identified seven dominant profiles (Fig. 2), which encompassed 66% of all isolates tested. As many as 42% belonged to the PFGE-pattern STYMXB.0131 (Fig. 2, identification code A), 13% to STYMXB.0079 (code B), and 4% to STYMXB.0010 (code C) (Hopkins et al., 2010). Patterns identified as codes D–G were found at rates below 3%. The frequency of STYMXB.0131 among the annual human serovar 4,[5],12:i:- isolates tested was 8.3% (95% CI, 2.3; 20.0) (4/48) in 2007 and reached a peak of 47% (95% CI, 39.3; 54.9) (79/168) in the year 2009. In contrast, the human STYMXB.0079 isolates represented 20.8% (95% CI, 10.5; 35.0) (10/48) of all human serovar 4,[5],12:i:- isolates in 2007 and decreased to 9.5% (95% CI, 5.5; 15.0) (16/168) in 2009. The human STYMXB.0010 isolates were found at a rate of 25% (12/48) in 2007 and decreased to 0.6% (95% CI, 0.01; 3.3] (1/168) in 2009. Patterns belonging to codes D–G showed no clear trend during this time period. It is worth mentioning that a considerable proportion, 118 of the 315 human ASSuT isolates (38%), was identified as STYMXB.0131, rendering this pulsotype the most frequent among the human isolates with the ASSuT resistance pattern. STYMXB.0131 strains in combination with this R-type were also frequently isolated in France, Germany, England, and Wales (Bone et al., 2010; Hauser et al., 2010; Hopkins et al., 2010). Interestingly, STYMXB.0131 was also the most prevalent pulsotype (60 of 71) among the monophasic pork isolates (Fig. 2). Moreover, 11 of the 22 poultry isolates belonged to this pulsotype (data not shown). In contrast to the human isolates—but in accordance with the pork isolates—the majority (7/11) of these poultry isolates expressed the SSuT resistance type, compared to only 2/11 showing the ASSuT. The Salmonella serovar 4,[5],12:i:- clone (PFGE pattern STYMXB.0131, resistance type SSuT) was isolated from humans as well as from pork/poultry meat, which gave strong evidence for an epidemiological link between pork/poultry meat and human disease. Of the 96 isolates belonging to the pulsotype STYMXB.0079, 52 corresponded to the ASSuT, and two to the SSuT resistance pattern. This pulsotype in combination with the R-type ASSuT was predominantly isolated from humans in Italy between 2003 and 2006 but not in other European countries (Dionisi et al., 2009; Hopkins et al., 2010). We speculate that this type was previously predominant in Switzerland and was replaced during the last several years by pulsotype STYMXB.0131.

Dominant pulsed-field gel electrophoresis patterns (A–G) of Salmonella enterica subsp. enterica serovar 4,[5],12:i:- during the years 2007–2011 in Switzerland. Profiles D, F, and G could not be assigned with reference strains of CDC PulseNet database. This figure shows their next relatives (STYMXB.0080, STYMXB.0206, STYMXB.0330, STYMXB.0055, STYMXB.0301, STYMXB.0199). Salmonella Braenderup H9812 was used as reference strain. Clustering was based on the unweighted pair-group method with averages, setting tolerance at 1% and optimization at 1.5%.
Based on the seven most common PFGE patterns found, 40 isolates were selected for further characterization with MLVA and phage typing. The main focus was made on type STYMXB.0131, of which 15 isolates of different origins, dates of isolation, and resistance patterns were selected. From the second most frequently encountered PFGE pattern—STYMXB.0079—we chose 10 different isolates: from STYMXB.0010 seven isolates and from the remaining four patterns two isolates each.
Phage typing showed 10 different types with the following distribution: 55% (22/40) belonged to DT193, 15% (6/40) to DT104blow, 12.5% (5/40) were RDNC (reaction pattern did not conform to the phage scheme), 10% (4/40) to U311, 5% (2/40) to DT120, and 2.5% (1/40) to DT7. Among the RDNC patterns, no commonality was found; therefore, they were counted as five different types. Again, these findings supported those reported earlier with the European 4,[5],12:i:- by Hopkins et al. (2010, 2012).
MLVA typing showed 13 different allele combinations (MLVA profiles), all of them with the same number of tandem repeats at locus STTR9 and with absence of the virulence plasmid locus STTR10 (Table 2). Differences were found at loci STTR5, STTR6, and STTR3. Irrespective of the PFGE profile, resistance pattern, or phage type, the MLVA profile 3-12-10-0-0211 predominated with a percentage of 35% (14/40). As illustrated in Table 2, profile 3-13-9-0-0211 appeared in 15% (6/40) of the isolates, and no more than 10% of the isolates could be assigned to any other MLVA profile. Only one isolate with profile 3-14-9-0-0111 differed from the others at locus STTR3. Locus STTR3 with allele no. 0211 and locus STTR9 with allele no. 3 were also typically found in other European serovar 4,[5],12:i:- isolates and, similarly, locus STTR10 was consistently absent, indicating the lack of the typical Salmonella Typhimurium virulence plasmid in those isolates (Hauser et al., 2010; Hopkins et al., 2010). Loci STTR5, STTR6, and STTR9 appear to be highly variable among the European and Swiss isolates.
STTR 9-5-6-10-3.
Human origin.
Food origin (pork).
Reaction pattern did not conform to the phage scheme.
Food origin (poultry).
Environmental origin.
MLVA, multilocus variable-number tandem-repeat analysis; ASSuT, ampicillin–streptomycin–sulfonamide–tetracycline resistance pattern; SSuT, streptomycin–sulfamethoxazole–tetracycline resistance pattern; SuT, sulfamethoxazole–tetracycline resistance pattern; TMP, trimethoprim; G, gentamicin; C, chloramphenicol.
Conclusions
In conclusion, XbaI PFGE analysis of all isolates revealed a large variety of profiles within the monophasic isolates. Nevertheless, among these we identified seven dominant profiles, which encompassed 66% of all isolates tested. Moreover, our data provide strong evidence for a spread of two specific Salmonella serovar 4,[5],12:i:- clones (PFGE pattern STYMXB.0131, resistance type ASSuT) and (PFGE pattern STYMXB.0131, resistance type SSuT) during these years in Switzerland. In contrast to the human isolates, the pork/poultry isolates expressed predominantly the SSuT resistance type. The Salmonella serovar 4,[5],12:i:- clone (PFGE pattern STYMXB.0131, resistance type SSuT) was isolated from humans as well as from pork/poultry meat, which gives strong evidence for an epidemiological link between pork/poultry meat and human disease. Studies such as the one presented here are able to provide sufficient scientific data to act as grounds on which to base law-enforcement activities by food safety authorities. For example, the fact that an important percentage of the human morbidity is caused by one particular clone found in a particular and traceable product justifies such action. Nevertheless, the limitations are also clearly shown, in that other clones also contribute to morbidity. Hence, precise studies help in fine-tuning legal actions.
Footnotes
Acknowledgments
We thank Helga Abgottspon, Simon Bohli, Grethe Sägesser for their help in strain collection and for their technical support, Ernst Junker for his help with MLVA, and Katie Hopkins and Tansy Peters from CDC PulseNet for their support with comparison of PFGE patterns with the PulseNet database. This work was financially supported by the Swiss Federal Office of Public Health and by the Federal Institute for Risk Assessment, Germany, BFR46-003.
Disclosure Statement
No competing financial interests exist.