
Abstract
The resistance of foodborne Staphylococcus aureus isolates to antimicrobial agents is an increasing problem. In this study, 154 S. aureus food isolates were examined on the basis of inducible clindamycin resistance and the presence of ermABC genes. Furthermore, the genetic relatedness of S. aureus strains having inducible and constitutive clindamycin resistance was investigated by pulsed-field gel electrophoresis. Inducible clindamycin resistance was determined in five strains using the D-test. Only S. aureus strain HE7A showed constitutive resistance. Polymerase chain reaction results demonstrated that all constitutive and inducible clindamycin resistant strains contained only the ermC gene. However, the presence of the ermC gene was also detected in three strains without inducible or constitutive resistance. Nucleotide variations on the sequences of ermC genes were also determined. Pulsed-field gel electrophoresis analysis of two groups of S. aureus strains (S4BY, PY62A, PY38BY/S and S35A, S35B) with inducible resistance demonstrated 100% homology, but the other susceptible and ermC gene carrying S. aureus strains showed different band patterns with 44–88% homology. This is the first study reporting inducible clindamycin resistance in foodborne S. aureus isolates in Turkey.
Introduction
T
Resistance to macrolides and lincosamides is common in S. aureus, but resistance to streptogramins remains infrequent (Lina et al., 1999; Fridkin et al., 2005). In staphylococci, three types of mechanisms are responsible for acquired resistance to MLS antibiotics: target site modification, active efflux, and drug modification. The predominant resistance mechanism is methylation, especially A2058/A2059 residues in domain V of bacterial 23S rRNA (Vandendriessche et al., 2011). Erm genes (mainly ermA, ermB, and ermC) encode erythromycin ribosome methylase, leading to MLS type B (MLSB) resistance phenotype (Leclercq and Courvalin, 1991). The resistance mechanism is either inducible (strains are resistant to 14- and 15-membered ring macrolides and susceptible to 16-membered ring MLSB) or constitutive (strains are also resistant to 16-membered ring MLSB) (Lina et al., 1999). Resistance to lincosamides is caused by methylation of bacterial 23S ribosomal RNA (Spížek and Řezanka, 2004). In staphylococci, resistance to streptogramin A and B is always associated with the presence of streptogramin A resistance determinants. When those determinants were combined with streptogramin B resistance determinants, for example erm gene (MLSB resistance phenotype) and vgb gene (SB phenotype), high-level synergic resistance was obtained (Cocito et al., 1997).
The aim of this study was to determine an inducible MLS resistance phenotype by the D-test, MLS resistance genes using polymerase chain reaction (PCR) assays, and genetic relatedness of resistant and related strains by pulsed-field gel electrophoresis (PFGE) analysis in previously isolated foodborne S. aureus strains.
Materials and Methods
Bacterial isolates and D-test
A total of 1070 food samples (452 dairy products, 303 raw milk, 141 bakery products, 130 meat and meat products, and 44 ready-to-eat foods) were collected from seven cities (Balikesir, Bursa, Canakkale, Edirne, Istanbul, Kirklareli, and Tekirdag) in the Marmara Region of Turkey from July 2007 to December 2008, and 154 S. aureus strains were previously isolated (Aydin et al., 2011). Isolation of S. aureus from food samples was performed using EN ISO 6881-1 standard procedures (ISO, 1999), and methods for the identification of isolates were previously described (Aydin et al., 2011). The D-test was carried out to determine inducible clindamycin resistance phenotype using erythromycin (15 μg) and clindamycin discs (2 μg) (Oxoid, Hampshire, UK) (Levin et al., 2005; CLSI, 2012). For this purpose, the strains were grown on Tryptic Soy Agar (Merck, Darmstadt, Germany) at 37°C overnight and the colonies were suspended in 10 mL 0.9% (wt/vol) NaCl. Bacterial suspensions were adjusted to McFarland 0.5 using a densitometer (Den-1, HVD Life Sciences, Austria) and inoculated onto Mueller-Hinton Agar plates (Oxoid), and then erythromycin and clindamycin discs were placed 17 mm apart (edge to edge). The plates were incubated at 35°C for 18 h, and the production of D-shaped zones between erythromycin and clindamycin discs was evaluated visually.
Detection of ermABC genes
Genomic DNA of bacterial strains was extracted as described previously (Sudagidan et al., 2008). The screening of MLS resistance determinant genes ermA (421 bp), ermB (359 bp), and ermC (572 bp) was performed by PCR using the primers designed by Lina et al. (1999). PCR reactions were carried out in a 50-μL reaction mixture containing 1.2 U Taq DNA polymerase (Fermentas, Vilnius, Lithuania), 5 μL of 10× reaction buffer (750 mM Tris-HCl (pH 8.8), 200 mM (NH4)2SO4, 0.1% Tween 20, and 1.5 mM MgCl2), 10 μM of each of the primers (Integrated DNA Technologies, Coralville, IA), 0.2 mM each of the four dNTPs (Fermentas), and 5 μL of genomic DNA as a template. PCR conditions were 10 min at 94°C, 35 cycles of 30 s at 94°C, 30 s at 52°C (for ermA and ermC) and at 55°C (for ermB) and 1 min at 72°C, and finally 10 min at 72°C. Then, PCR products were resolved in 1.5% (wt/vol) agarose gel in 1× TAE buffer. PCR experiments were repeated at least twice for each strain. The obtained PCR amplicons were sequenced in Applied Biosystems 3130xl Genetic Analyzer (Carlsbad, CA) directly by the same primers used in PCR.
PFGE
Genetic relatedness of inducible/constitutive clindamycin-resistant and related S. aureus strains was determined by PFGE analysis. Agarose plugs were prepared as described previously (Durmaz et al., 2007). Bacterial DNA in agarose plugs was digested with 30U SmaI (Fermentas) at 30°C overnight and they were run in 1% (wt/vol) pulsed-field certified agarose (Bio-Rad, Hercules, CA) with 5-40-s pulse time, 6 V/cm, 120° angle, at 14°C for 22 h using CHEF-Mapper PFGE system (Bio-Rad). After electrophoresis, the gel was stained with 0.5 μg/mL ethidium bromide and visualized in VersaDoc 4000MP image analyzer (Bio-Rad). PFGE band patterns were analyzed using BioNumerics software version 6.6 (Applied Maths, Belgium), and the similarity among PFGE band patterns was evaluated using unweighted pair group method with arithmetic mean similarity coefficient with 1% band tolerance and 0.5% optimization.
Results
A total of 154 isolated S. aureus strains were analyzed and 3.2% S. aureus strains (S35A, S35B, S4BY, PY62A, and PY38BY/S) displayed inducible clindamycin-resistance phenotype, and only S. aureus strain HE7A showed constitutive resistance; in other words, it was resistant to both erythromycin and clindamycin. All inducible and constitutive clindamycin-resistant strains were positive for only the ermC gene. Although S. aureus strains PY4, PY5, and PY178B were susceptible to both erythromycin and clindamycin (Aydin et al., 2011), they were also found to contain the ermC gene. DNA sequencing of ermC PCR amplicons demonstrated that the nucleotide sequence variations were present (Table 1). The dendrogram shows the results of PFGE analysis (Fig. 1). Three resistant strains (S4BY, PY62A, and PY38BY/S) belonging to the same cluster were isolated in the Kesan region. However, they were isolated from three different food sources. Additionally, S35A and S35B with inducible resistance showed 100% homology with indistinguishable band patterns. However, the constitutive resistant strain HE7A was unrelated to other susceptible and inducible resistant strains. Erythromycin- and clindamycin-susceptible strains (HE4D, HE25A, HE25B, HE27A, PY67A, PY30AY/B, PY30AY/S, and PY38BY/B) isolated in the Kesan region and ermC carrying susceptible strains (PY4, PY5, and PY178B) showed 44–88% homology, and they were not closely related to the resistant strains (Fig. 1).
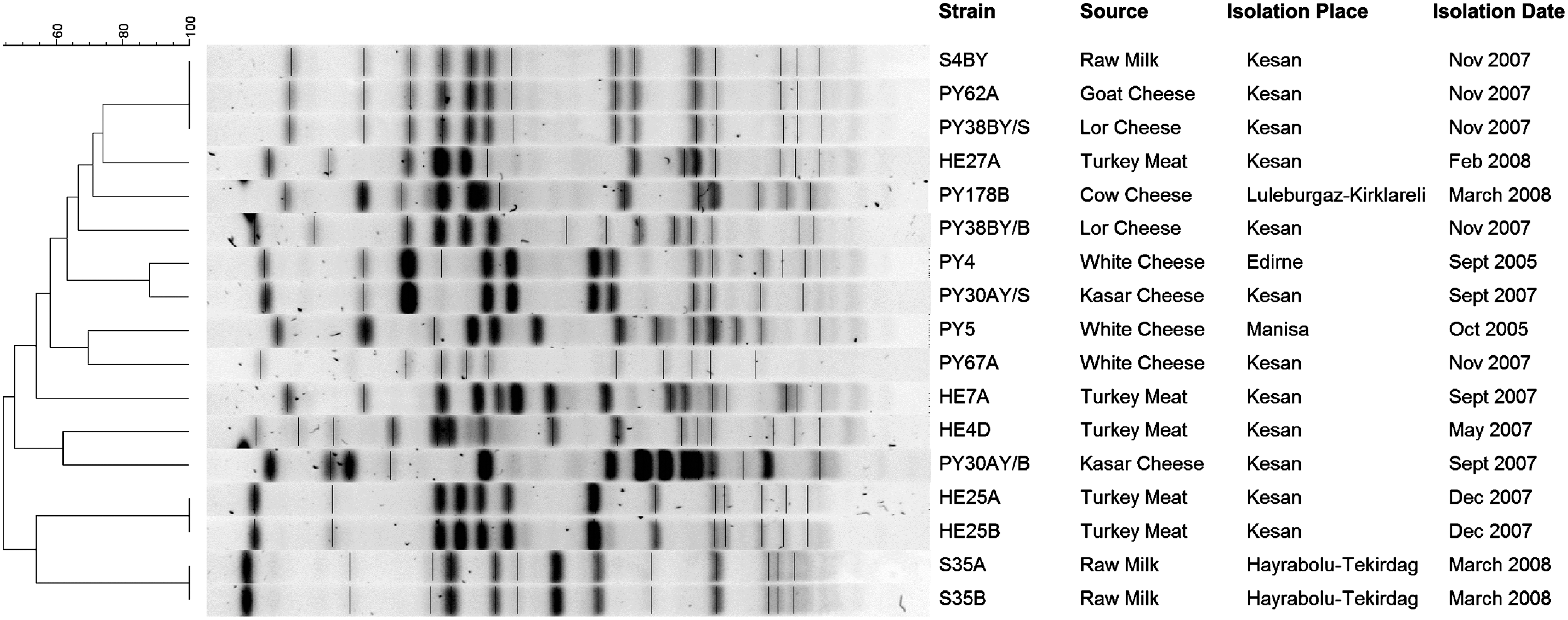
Restriction band patterns obtained by pulsed-field gel electrophoresis analysis indicated the genetic relatedness of Staphylococcus aureus strains with inducible/constitutive clindamycin resistance, ermC gene–positive susceptible strains, and the susceptible strains isolated in the Kesan region.
Discussion
The increased use of antibiotics both in the treatment and feed of animals as food additives can cause an increase in the survival of resistant bacteria. S. aureus is one of the important causative agents in foodborne intoxications, mastitis, scalded-skin syndrome, and Panton-Valentine Leukocidin–induced necrotizing pneumonia. Transmission of resistant and virulent S. aureus strains from humans to food or from food to humans leads to spread of these strains. In this study, a low prevalence of inducible and constitutive clindamycin-resistance phenotypes were found among 154 S. aureus strains. In addition, six resistant and three susceptible strains contained only the ermC gene. The nucleotide variations on ermC DNA sequences of amplified PCR products were also found with no specific nucleotide changes for inducible/constitutive or susceptible phenotype. However, ermC-gene-carrying susceptible strains (PY4 and PY5) were resistant only to penicillin G and strain PY178B was resistant only to tetracycline (Aydin et al., 2011). MLS-resistant strains were multidrug resistant to more than three antibiotics, especially penicillin G, kanamycin, and tetracycline, whereas all of them were methicillin-susceptible S. aureus (Aydin et al., 2011). In another study, a higher inducible MLS resistance phenotype was detected in 52.8% (38/72) of S. aureus isolates from cows with clinical mastitis in China, and the most dominant resistance gene was ermC (59/67; 88.1%), followed by ermB (16/67; 23.9%) among erythromycin-resistant strains (Wang et al., 2008). On the other hand, Ochoa-Zarzosa et al. (2008) isolated 31 S. aureus strains from 106 raw milk composite samples of cows with subclinical mastitis, and they obtained two erythromycin-resistant strains (STA9 and STA13) containing ermB and ermC genes, respectively. Additionally, Türkyılmaz et al. (2010) isolated 93 S. aureus strains from bovine milk with mastitis in Turkey and they found nine ermA and seven both ermA- and ermB-positive strains among macrolide-resistant strains. However, none of them was positive for the ermC gene. The predominant gene was ermC in 51 of 55 inducible MLS-resistant strains, whereas in oxacillin-resistant S. aureus strains, the ermA gene was mostly detected (27 of 29) in both mastitis cases and in clinical oxacillin-susceptible S. aureus strains (Janapatla et al., 2007). In Denmark, the prevalence of erm genes in S. aureus isolates from blood cultures indicated that ermC gene was present on plasmids and the predominant one was 2.5 kb in size in 77 isolates (Westh et al., 1995). In this point of view, the presence of the ermC gene in MLS-susceptible strains could be explained by carrying related plasmids. Additionally, Azap et al. (2005) reported the presence of inducible clindamycin-resistant clinical staphylococcal isolates in Turkey. In that study, 408 isolates were tested and inducible clindamycin resistance was detected in 5.7% of 105 methicillin-resistant S. aureus isolates, 3.6% of 111 methicillin-susceptible S. aureus isolates, 30.8% of 94 methicillin-resistant coagulase negative staphylococci (CNS) isolates, and 11.2% of 98 methicillin-susceptible CNS isolates.
On the basis of PFGE results, three inducible resistant strains from different sources showed 100% homology. However, constitutive strain HE7A had band patterns that were distinct from the susceptible strains and the strains showing inducible clindamycin resistance. The susceptible strains isolated in the Kesan region showed different PFGE band patterns, and they were unrelated to resistant strains isolated in the Kesan region (Fig. 1). These results suggest that there is no unique clonal type that shows inducible resistance to clindamycin.
It could be concluded that the presence of inducible clindamycin-resistant and multidrug-resistant strains are in low prevalence among foodborne S. aureus strains. More analysis is needed on foodborne S. aureus strains from different countries to determine the prevalence of inducible clindamycin resistance in food. The sources of these strains and resistance mechanisms other than erm genes or the role of nucleotide changes remain to be elucidated.
Footnotes
Acknowledgments
The authors thank Izmir Institute of Technology, Biotechnology and Bioengineering Applications and Research Center (BIYOMER) for PFGE analysis and Dane Ruscuklu for DNA sequencing studies. The authors also thank David Ames Mann, Research Professional I (University of Georgia, Center for Food Safety) for critical reading of the manuscript. This study was supported by The Scientific and Technological Research Council of Turkey (TUBITAK) (Project No: 107T266) and by the Research Fund of Istanbul University, Project No: UDP-19101/2011.
Disclosure Statement
No competing financial interests exist.