
Abstract
Weaned piglets are very susceptible to diarrhea caused by enterotoxigenic Escherichia coli. In the past, various natural components were proposed to have beneficial effects by reducing the effects of diarrheal infectious diseases in humans and animals, and thus may represent an alternative for the use of (prophylactic) antibiotics. Alternatives may inactivate enterotoxigenic Escherichia coli heat-labile toxin (LT) by interfering with toxin binding to the cellular receptor GM1. In this study, various plants and other natural substances were tested for inhibitory properties, in the GM1 binding assay, and in the LT-induced cAMP production in Vero cells. The toxic dose of each compound was determined in a cell viability assay, and the highest nontoxic concentrations were used in the GM1 and cAMP assays. Results demonstrated that only
Introduction
Blocking the receptor
Lectins are sugar-binding proteins that are highly specific for their specific sugar moieties. Several lectins are known to bind to the galactose and N-acetyl-
Competing LT binding sites
Galactose-containing or -related oligosaccharides
Because galactose is so important for toxin receptor binding recognition, it is hypothesized that galactose-like sugars could be an alternative binding site for the toxin instead of the GM1 ganglioside (Fukuta et al., 1988; Holmner et al., 2011).
Yeast cells and yeast cell extracts
These cells and extracts contain components such as manno-oligosaccharides, which could bind enterotoxin. The nonpathogenic yeast Saccharomyces boulardii is used in many countries for the treatment of gastrointestinal disorders (Schellenberg et al., 1994).
Direct inactivation
Tea extracts are traditional herbal medications used in China and Japan for many centuries to treat diarrhea. They contain polyphenols, which can form aggregates with enterotoxin, rendering them inactive (Verhelst et al., 2010).
Here, several natural products from each group were tested for activity in a GM1 binding assay, and for the capacity to reduce LT-induced cAMP accumulation in the Vero cell line. To exclude aspecific toxic effects, products were first tested for their toxicity in the Vero cell line; the highest nontoxic concentration or a 2 mg/mL concentration was used for toxin inactivation in both assays.
Materials and Methods
Products
GM1 ganglioside,
GM1-enzyme-linked immunosorbent assay (ELISA)
All steps were performed at room temperature. Twenty-five microliters of GM1 in ethanol (10 μg/mL) was added to each well of a 96-well plate. Thereafter, the plate was incubated for 2 h. All unattached GM1 was removed by washing the plate three times with phosphate-buffered saline Tween (PBST, pH 7.2, 0.010 mg/ml Tween) and the remaining binding sites of the surface were blocked for 1 hour with Synblock followed by washing with PBST. Then 50 μL of heat-labile toxin (0.5 μg/mL) with 50 μL of a test compound was added. After 1-h incubation, the unbound toxins were removed by washing three times with PBST. Primary anti-LT antibodies (100 μL, 1:500) were added to each well and incubated for 1 h. Before the secondary antibodies (rabbit anti-immunoglobulin G) (100 μL, 1:1000) were added, the plate was washed three times with PBST. After 1 h, the ABTS substrate (100 μL) was added to each well and absorbance was measured at 405 nm (Morinaga et al., 2005).
Vero cell culture
Stock cultures of Vero cells (ATCC) were grown in Eagle's MEM medium supplemented with 10% fetal calf serum (FCS) in 6% CO2 at 37°C and were trypsinized and centrifugated (400×g) for 5 min. Media was removed and cells were counted and diluted to a concentration of 75,000 cells/mL medium. In a 96-well plate, 100 μL of cells was added to each well and left for incubation at 37°C overnight, before use in either the cell viability assay or the cAMP assay.
Cell viability assay (MTT assay)
Vero cells were treated with serially twofold-diluted compounds starting from 10 mg/mL in culture medium. After 24 h, 20 μL of 5 mg/mL MTT was added to each well and left for incubation for 3.5 h at 37°C. Living cells reduce yellow MTT to insoluble purple formazan. The latter is solubilized by adding 150 μL of MTT solvent by shaking for 15 min. Absorbance was read at 590 nm with a reference filter of 620 nm (Mossmann, 1983).
cAMP assay
The natural components (at the noncytotoxic concentrations indicated in the results section) were incubated with LT (50 nM, in phosphate-buffered saline pH 7.2) to evaluate reduction of LT-induced cAMP accumulation with phosphate-buffered saline as a negative control. These mixtures (100 μL) were then added for 20 min to the Vero cells and cAMP accumulation was quantified using a cAMP-Glo Assay (Promega, Madison, WI) according to the manufacturer's instructions.
Statistics
Statistical software program Graphpad Prism 5.0 for Windows (Graphpad Software, San Diego, CA) was used to carry out all statistics. The data sets did not follow a normal distribution according to the Kolmogorov-Smirnov normality test. Therefore, the nonparametric Mann–Whitney U test of significance (significance level was set p<0.05) was used to compare values of treatments versus control.
Results
Determining the highest nontoxic concentration
A wide range of natural components concentrations was screened to verify if they could affect Vero cell viability. Among those screened, the highest nontoxic concentration of each natural component (defined as MTT values higher than 75% survival) was subsequently used to investigate the protective activity against LT binding and LT-induced cAMP production. Yeast and yeast extract appeared to have no toxicity in this range, and a 2 mg/mL concentration was used for toxin inactivation in both assays. The highest nontoxic concentration was for galactose, N-acetyl-
Evaluation of natural components inhibiting LT binding to GM1
LT (0.5 μg/mL) was preincubated for 60 min before being applied to the GM1-ELISA. The data indicate that some but not all natural compounds were able to prevent the binding of LT to its receptor GM1.
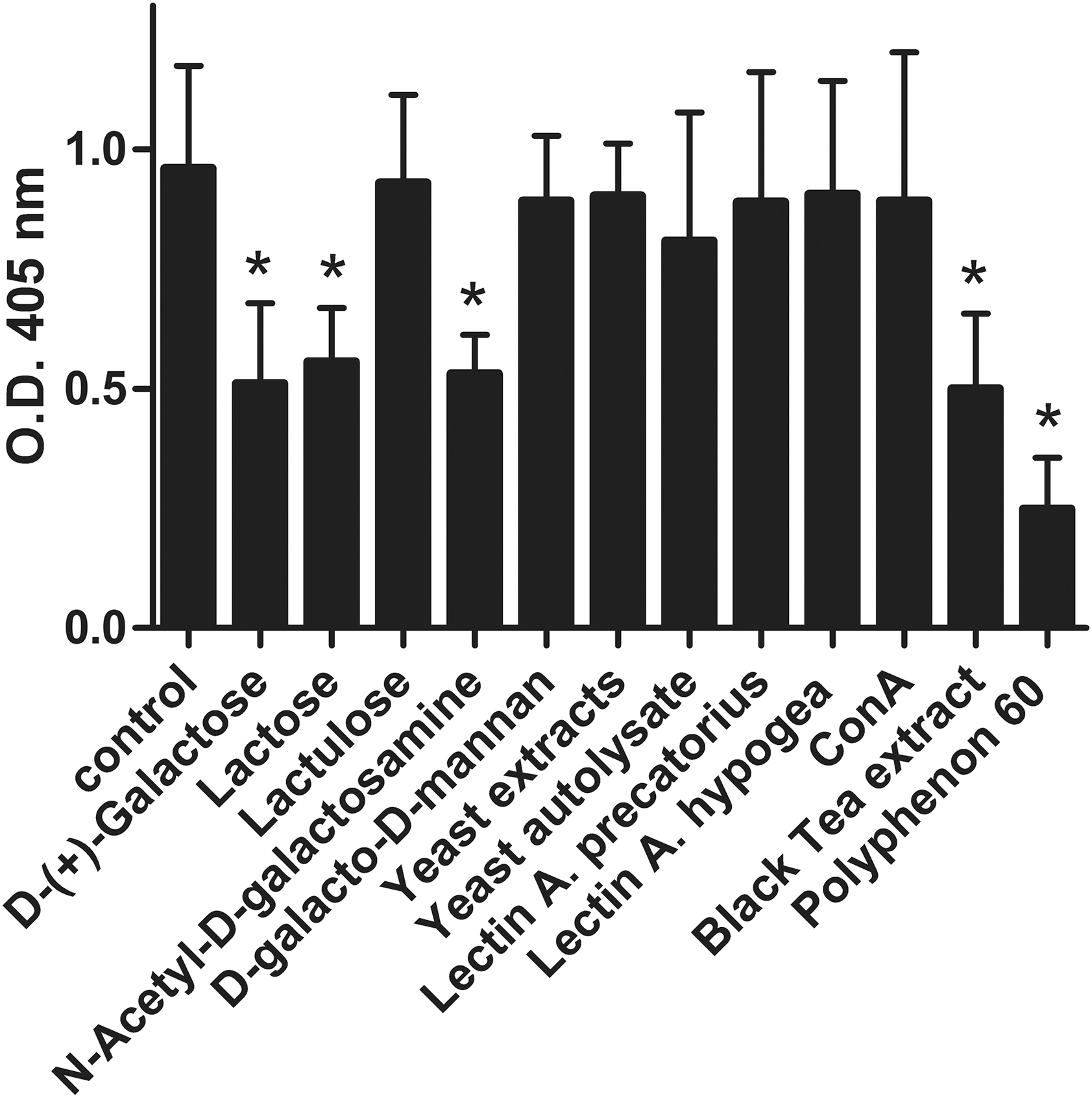
Significant inhibition of heat-labile toxin binding to GM1 by
Effect of active compounds on LT-induced cAMP concentration
Because the above results suggested that several natural components were able to prevent the binding of LT to its receptor, we wanted to assess whether they are also capable of preventing the LT-induced cAMP production in Vero cells. In Vero cells, co-incubation of LT with the active natural compounds revealed that extract from black tea and Polyphenon 60 from green tea were able to significantly lower the cAMP concentration to 56% and 61% of the control, respectively.
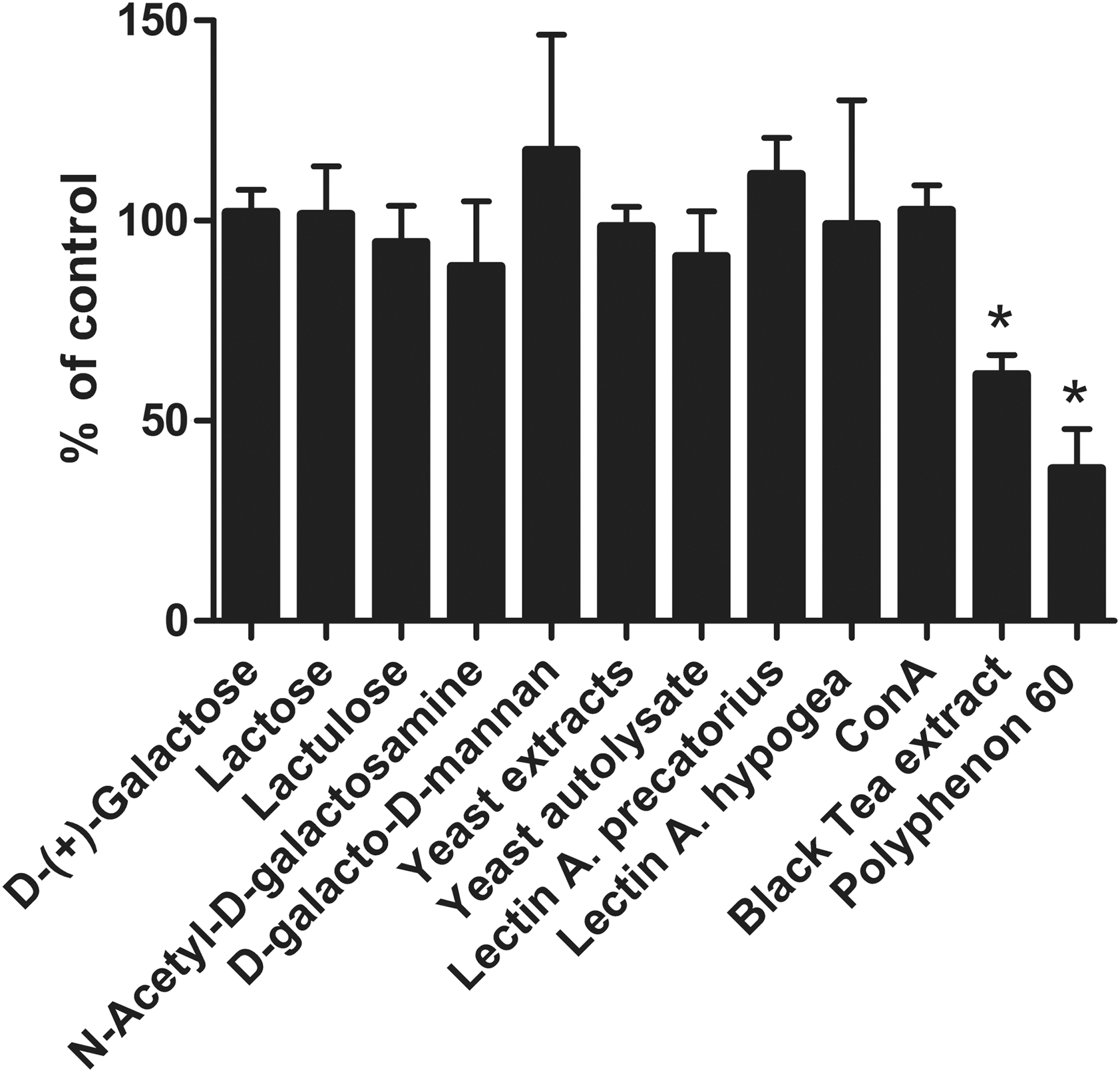
Significant inhibition of heat-labile toxin (LT)–induced cyclic adenosyl monophosphate (cAMP) concentration in Vero cells by black tea extract, and Polyphenon 60. Data are expressed as a percentage of the cAMP level induced by LT alone in the control. Data are means±standard error of the mean of values of three experiments. Significant differences (p<0.05) from control are indicated with an asterisk. Con A, concanavalin A.
Discussion
Because LT is the major virulence factor in ETEC diarrhea, natural inhibitors of LT could show promise as an in-feed alternative for antibiotics. Our results show that black tea extract, Polyphenon 60, galactose, N-acetyl-
Based on the results of the GM1 ELISA, a similar result was anticipated in the Vero cell cAMP assay. This was true for black tea extract and Polyphenon 60. However, to our surprise, galactose, galactosamine, and lactose did not lower LT-induced cAMP production in the Vero cell assay. The most likely explanation is that a large part of these compounds was quickly metabolized in the cell culture, and at least galactose was described to be rapidly metabolized by Vero cells (Mendonça et al., 2002). Because the same may happen in vivo by intestinal cells, this means that either it is very unlikely that these compounds are active in vivo, or that in order to maintain activity the already very high concentrations used here should even be increased. The only natural compounds that were able to both prevent the binding of LT to GM1 and to lower cAMP concentration in Vero cells were the tea extracts. The chewing of tea leaves is well known as a folk medicine to reduce diarrhoea. Tea leaves are characterized by high concentrations of polyphenols (Bruins et al., 2006). Previous research in intestinal loops of mice demonstrated that catechins and black tea extract were able to reduce ETEC-induced fluid secretion in an animal model (Bruins et al., 2006). Polyphenols have been reported to inhibit the binding of CT to GM1 and also reduce CT-caused cAMP production in Vero cells through aggregation (Saito et al., 2002; Morinaga et al., 2005; Morinaga et al., 2010). Using the same system with LT and purified polyphenols from tea, we recently demonstrated that only polyphenols with two or more galloyl moieties in the molecule are responsible for the toxin-inactivating properties (Verhelst et al., 2013). Furthermore, polyphenols have a low absorption in the small intestine (Biasi et al., 2011). The latter fact, and the lower effective concentrations means that polyphenols can be used at realistic and commercially feasible inclusion levels in feed.
In conclusion, our data show that combining the GM1 ELISA with the Vero cell assay is a powerful method for selecting effective inhibitors, because it includes a cell culture that better mimics the in vivo situation. It also shows that blocking of LT-receptor interactions forms a promising avenue for therapeutic intervention and can lead to a wide new range of natural products to be used as an alternative for in-feed antibiotics.
Footnotes
Acknowledgments
This work was carried out within the framework of the IWT project 60680 sponsored by the Flemish Institute for the Promotion of Scientific and Technological Research in the Industry (IWT).
Disclosure Statement
No competing financial interests exist.