
Abstract
Non-O157 Shiga toxin–producing Escherichia coli (non-O157 STEC) strains are increasingly recognized as important foodborne pathogens worldwide. Together with E. coli O157:H7, six additional STEC serogroups (O26, O45, O103, O111, O121, and O145) are now regulated as adulterants in certain raw beef products in the United States. However, effective detection and isolation of non-O157 STEC strains from food matrices remain challenging. In the past decade, great attention has been paid to developing rapid and reliable detection methods for STEC in general (targeting common virulence factors) and specific STEC serogroups in particular (targeting serogroup-specific traits). This review summarizes current trends in detecting non-O157 STEC in food, including culture, immunological, and molecular methods, as well as several novel technologies.
Introduction
A
On the other hand, non-O157 STEC strains are increasingly recognized as important foodborne pathogens worldwide, with reported outbreaks linking to produce, milk, juice, and beef, among others (Eblen, 2007; Mathusa et al., 2010; Grant et al., 2011; CDC, 2012b; USDA, 2012b). Although non-O157 STEC strains appear to cause watery diarrhea more often than bloody diarrhea, in certain geographic regions (e.g., Latin American, Australia, and Europe), the frequency of non-O157 STEC implicated in hemolytic–uremic syndrome cases actually rivaled that of E. coli O157:H7 (Karmali et al., 2003; Johnson et al., 2006). Reporting of non-O157 STEC in the United States has increased every year since it was designated a nationally notifiable infection in 2000 (Atkinson et al., 2006). Recent FoodNet data suggest that non-O157 STEC infections have started to gain predominance over O157 cases in the United States (CDC, 2011, 2012a, 2013). Six STEC serogroups—O26, O45, O103, O111, O121, and O145 (referred to as “top 6” hereafter) —accounted for approximately 75% of total non-O157 STEC illnesses in the United States annually (Brooks et al., 2005; USDA, 2012b), while other highly pathogenic serogroups (e.g., O91, O104, O113, and O128) appear more prevalent in countries outside of the United States (Karmali et al., 2003; Johnson et al., 2006; Bettelheim, 2007; EFSA/ECDC, 2012).
Considering the rising public health concern about non-O157 STEC, effective June 4, 2012, FSIS expanded the zero-tolerance policy for E. coli O157:H7 to include the top 6 non-O157 STEC serogroups in raw, nonintact beef products (USDA, 2012c). With this regulation, it is imperative that rapid, accurate, and reliable detection methods be available to test for non-O157 STEC in beef and other high-risk foods. Strategic Consulting Inc. (
Challenges
There are many inherent challenges associated with pathogen detection in food, such as low levels of injured/stressed target cells, high levels of background flora, nonhomogeneous distribution of target pathogens, and complex food matrices involved (Ge and Meng, 2009). As a result, effective sampling and sample preparation prior to the actual analyses are deemed critical (Stevens and Jaykus, 2004; Brehm-Stecher et al., 2009; Dwivedi and Jaykus, 2011). Enrichment is often used to overcome these challenges and primarily serves to (1) resuscitate injured/stressed target cells; (2) increase the target cell numbers as much as a million-fold; and (3) dilute the effect of food inhibitors and background flora on the assay. However, with enrichment, the sample-to-result time is inevitably extended to days rather than hours (Ge and Meng, 2009).
Additional challenges unique to non-O157 STEC detection in food are evident. First, unlike E. coli O157:H7, phenotypic characteristics (e.g., sorbitol fermentation) that distinguish non-O157 STEC from generic E. coli are lacking. Therefore, non-O157 STEC is neither readily nor routinely cultured in clinical diagnostic and food-testing laboratories (Gould et al., 2009). Second, non-O157 STEC encompasses a diverse group of organisms with a total of 174 E. coli O-serogroups identified to date, although not all are capable of producing Shiga toxins (DebRoy et al., 2011a). Even highly pathogenic non-O157 STEC still comprises 4–10 serogroups (Karmali et al., 2003; Johnson et al., 2006; EFSA, 2009; USDA, 2012a), requiring a suite of assays for detection and confirmation. Third, a stepwise approach is commonly used for non-O157 STEC, first screening for virulence genes (e.g., Shiga toxin genes [stx] and the E. coli attaching and effacing gene [eae]), then testing for O serogroup–specific genes (USDA, 2012a). Due to the lack of unequivocal virulence markers, nonpathogenic STEC strains may also give signals for the target genes at the first-step screening. It is also challenging to confirm that the signals for stx, eae, and serogroup-specific genes in the enrichment samples are originated from individual cell or from a mixture of different cells; therefore, further confirmation steps are essential.
Overview of Method Developments
In the United States, food safety regulatory agencies such as the U.S. Food and Drug Administration (FDA) and USDA have each developed and validated a set of detection methods for foodborne pathogens of interest in various foods. Methods adopted for non-O157 STEC are available online at FDA's Bacteriological Analytical Manual (BAM) chapter 4A (FDA, 2012a) and FSIS's Microbiology Laboratory Guidebook (MLG) method 5B (USDA, 2012a). Schematic diagrams of the two methods are shown in Figure 1; the methods combine culture, immunological, and molecular techniques as described below and take about 4–5 days.

Schematic diagrams of current procedures employed by the U.S. Food and Drug Administration (FDA)
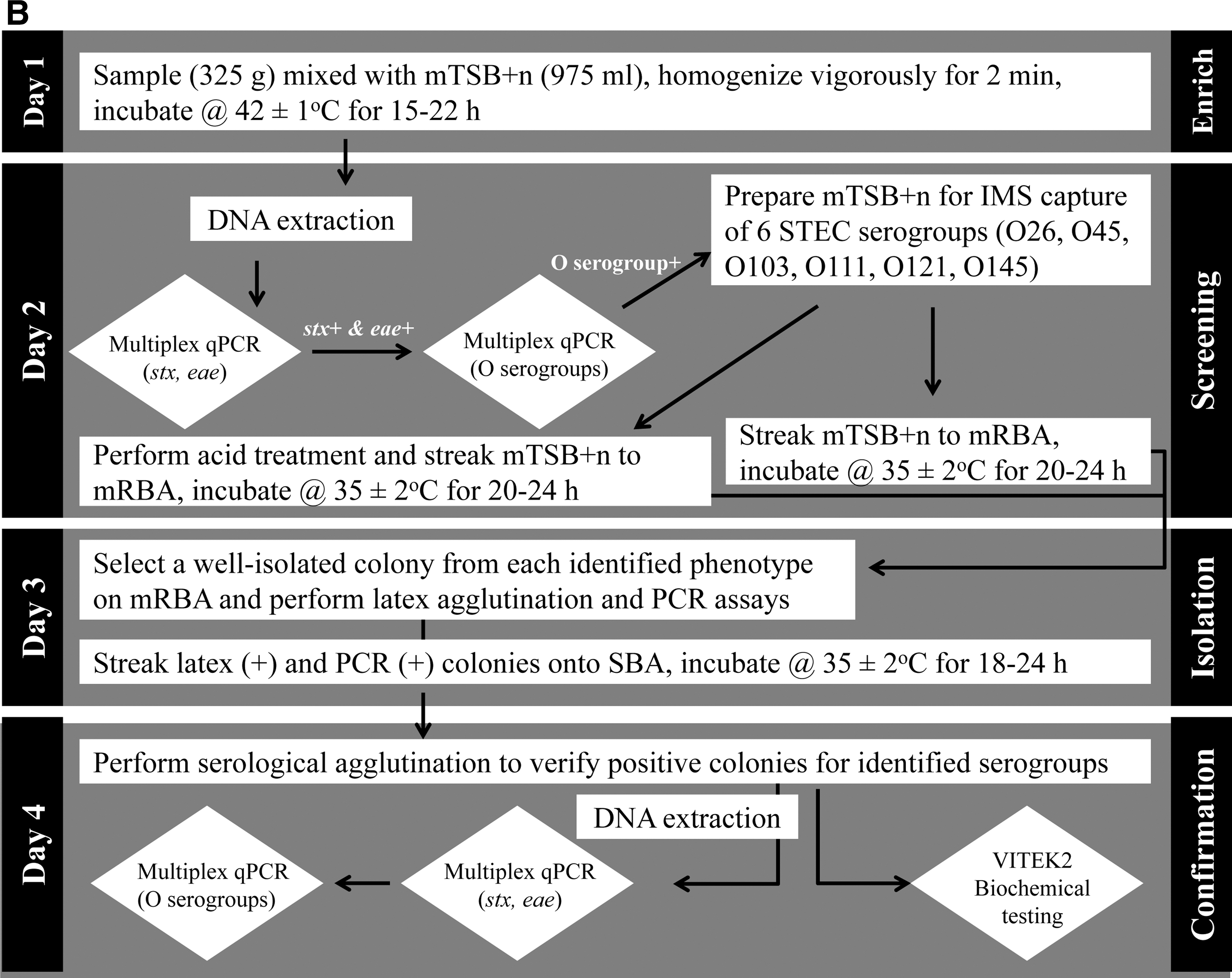
Many test kit manufacturing companies are also geared toward developing rapid detection methods for emerging pathogens such as non-O157 STEC. Table 1 includes a select list of commercially available non-O157 STEC test methods, several of which have recently received no-objection letters issued by FSIS (USDA, 2012d). Validation and certification of such methods are conducted by independent organizations such as the AOAC International, the International Organization for Standardization (ISO), and the Association French Normalization Organization Regulation. Very recently, ISO approved a real-time PCR-based method (ISO/TS 13136) for the detection of STEC O157, O111, O26, O103 and O145 in food and animal feed (ISO, 2012). Furthermore, many academic and government research laboratories are actively involved in method development for foodborne pathogens, including non-O157 STEC, driving the innovation to an even faster pace. Below current trends in non-O157 STEC detection, isolation, and confirmation in food are reviewed.
Top 6 non-O157 STEC serogroups consist of O26, O45, O103, O111, O121, O145.
The assays have been approved by the U.S. Food and Drug Administration's Center for Devices and Radiological Health for in vitro diagnosis of STEC infections.
The assays have been validated by the AOAC's Performance Tested Methods Program.
No-objection letters were issued by U.S. Department of Agriculture Food Safety and Inspection Service for these non-O157 STEC assays as of January 8, 2013 (USDA, 2012d).
Culture-Based Methods
Generally regarded as the criterion standard for pathogen detection in food, culture-based methods consist of multiple incubation steps (pre-enrichment, selective enrichment, selective and differential plating) for bacterial isolation followed by additional biological, serological, or molecular tests for confirmation (Feng, 2007; Ge and Meng, 2009). The hallmark of such methods is the use of suitable selective and differential culture media. Many of the media currently in use for non-O157 STEC are modified from those originally developed for E. coli O157:H7. For example, novobiocin is added to non-O157 enrichment broth at a lower concentration (e.g., 16 μg/mL) than the usual 20 μg/mL for E. coli O157:H7 due to less tolerance of non-O157 STEC to novobiocin (Vimont et al., 2007). Replacing sorbitol used in sorbitol MacConkey (SMAC) for culturing E. coli O157:H7 with rhamnose has led to the development of rhamnose-MacConkey (RMAC) for differential isolation of STEC O26 due to their inability to ferment rhamnose (Hiramatsu et al., 2002). Similar to E. coli O157:H7, tellurite-containing media have been used widely for selective culturing of STEC O26, O111, and O145 because of high-level tellurite resistance observed among these serogroups (Hiramatsu et al., 2002; Catarame et al., 2003; Orth et al., 2007). There are also several commercially available chromogenic agars for non-O157 STEC isolation, e.g., modified rainbow agar (mRBA; Biolog, Inc., Hayward, CA) (FDA, 2012a; USDA, 2012a) and CHROMagar STEC (CHROMagar Microbiology, Paris, France) (Hirvonen et al., 2012; Tzschoppe et al., 2012).
Recently, Possé medium based on a mixture of carbohydrate sources (sucrose and sorbose), β-D-galactosidase activity, and selective compounds (bile salts, novobiocin, and potassium tellurite) has been described for color-based differentiation of STEC O26, O103, O111, and O145. Confirmation of suspected colonies would require plating on additional agars containing dulcitol, rhamnose, raffinose, or arabinose (Possé et al., 2008). The isolation efficiency of Possé medium for low levels of non-O157 STEC in various dairy and meat products ranged from 64.3-100% (Possé et al., 2008). However, one independent evaluation of Possé medium in food applications reported ambiguity in the color differentiation between non-O157 STEC serogroups (Mathusa et al., 2010). Another recent development is the STEC heart infusion washed blood agar with mitomycin C (SHIBAM agar) (Lin et al., 2012) adopted by the FDA's BAM (Fig. 1). SHIBAM agar modifies washed sheep blood agar by adding mitomycin C and optimizes both the washed blood and base agar to facilitate STEC isolation (Beutin et al., 1996; Kimura et al., 1999; Sugiyama et al., 2001; Lin et al., 2012). Evaluation of SHIBAM agar showed that 89% (365 of 410) of STEC strains were hemolytic, while 86.3% (63 of 73) of non-STEC strains were not; SHIBAM also greatly improved the recovery of non-O157 STEC from artificially inoculated produce at<10 colony-forming units (CFU)/25 g when compared with Levine's eosin-methylene blue (L-EMB) agar (Lin et al., 2012).
For non-O157 enrichment, base media including trypticase soy broth (TSB), buffered peptone water (BPW), and E. coli broth (EC) are commonly used and frequently supplemented with novobiocin, cefixime, potassium tellurite, vancomycin, acriflavin, and cefsulodin to suppress the growth of background flora (Vimont et al., 2006; Hussein and Bollinger, 2008). The effects of enrichment broth, selective agent, and incubation protocol on the outcome of non-O157 STEC enrichment in various foods have been evaluated (Catarame et al., 2003; Vimont et al., 2006; Baylis, 2008; Hussein and Bollinger, 2008; Verstraete et al., 2010; Kanki et al., 2011; Gill et al., 2012). Catarame et al. (2003) reported that the optimum condition to recover E. coli O26 and O111 from minced beef was enrichment in TSB supplemented with cefixime (50 μg/L), vancomycin (40 μg/mL), and potassium tellurite (2.5 μg/mL; this compound was omitted for E. coli O111). In contrast, BPW was found to give increased recovery rates of both STEC O26 and O111 compared with TSB and EC broth (Drysdale et al., 2004). Baylis (2008) compared 10 enrichment broths and found non-O157 STEC strains only grew poorly in EC or modified EC (mEC) supplemented with novobiocin (20 μg/mL), likely due to the inhibitory effect of novobiocin at 20 μg/mL as mentioned above. More recently, Gill et al. (2012) examined six selective agents (cefixime, cefsulodin, mitomycin C, novobiocin, tellurite, and vancomycin) either singly or in combination for the optimal enrichment of a panel of 18 STEC serogroups in modified trypticase soy broth (mTSB), and found a combination of vancomycin (10 μg/mL) and cefsulodin (3 μg/mL) enabled the growth of all strains. Current FDA's BAM procedure for STEC enrichment in food uses modified BPW (containing casamino acids) with pyruvate for 5 h then supplemented with acriflavin (10 μg/mL), cefsulodin (10 μg/mL), and vancomycin (8 μg/mL) (FDA, 2012a), while USDA uses mTSB supplemented with novobiocin (8 μg/mL) and casamino acids (USDA, 2012a).
Several studies reported that incubating samples at 42°C was more effective than 37°C for isolating STEC O26, O111, and other STEC from various food samples ( Hara-Kudo et al., 2000; Catarame et al., 2003; Drysdale et al., 2004; Gill et al., 2012). We (Wang and others, unpublished data) also observed that 42°C enrichment was clearly more effective than 37°C for recovering low levels of the top 6 STEC serogroups from various produce items (lettuce, spinach, alfalfa sprouts). The current FDA's BAM protocol uses pre-enrichment at 37°C for 5 h to resuscitate injured/stressed cells followed by selective enrichment at 42°C overnight (FDA, 2012a), whereas the USDA method uses incubation at 42°C for 15–22 h (USDA, 2012a).
Recently, exposure to strong acidic conditions (TSB adjusted to pH 2 or 3) for a short period of time (30 min–2 h) was used to effectively reduce background flora before enrichment (Grant, 2005; Hu et al., 2009). The USDA method also applies a similar acid treatment step postenrichment to facilitate the isolation of non-O157 STEC strains from enrichment cultures (USDA, 2012a).
Immunological-Based Methods
Antibodies that recognize Shiga toxins or specific O antigens have been used in the direct detection of STEC serogroups in food samples or aid in their isolation through immunomagnetic separation (IMS) or colony immunoblot. Traditional cell cytotoxicity assays for the Vero and HeLa cell lines are very sensitive in detecting Shiga toxins but they are labor intensive, time consuming, and costly (Paton and Paton, 1998b; Johnson et al., 2006). Interestingly, simple and cost-effective Vero cell–based assays have been described recently (Zhao and Haslam, 2005; Quinones et al., 2009). Since first introduced to the United States in 1995, Shiga toxin enzyme immunoassays (EIAs) have gradually gained acceptance by clinical laboratories (Hoefer et al., 2011). To date, six Shiga toxin EIAs (Table 1) have been approved by the FDA for in vitro diagnosis of STEC infections (FDA, 2012b). Other commercially available EIAs include the Ridascreen Verotoxin EIA (R-Biopharm AG, Darmstadt, Germany) and VTEC-RPLA (Oxoid Ltd., Hampshire, United Kingdom). The time required to run these assays ranges from 20 min to 4 h, although overnight enrichment prior to the analysis is strongly recommended (Gould et al., 2009). Sensitivity of the EIAs can be enhanced by adding mitomycin C or polymyxin B in enrichment broth to stimulate the production or liberation of Shiga toxins (Johnson et al., 2006).
The application of Shiga toxin EIAs in food has been evaluated. Ridascreen and Premier EHEC (Meridian BioScience, Inc., Cincinnati, OH) were found to be 10-fold more sensitive than ProSpecT (Remel, Lenexa, KS); however, all three assays failed to detect Stx2d and Stx2e variants (Willford et al., 2009). Another study reported Premier EHEC to be useful for STEC screening in various food samples including raw milk, fresh and processed meat, and dairy food (Pontello et al., 2003). Very recently, Duopath Verotoxins kit (Merck, Darmstadt, Germany) was incorporated in a proposed streamlined method for STEC detection in food (Gill et al., 2012). Currently, Duopath Verotoxins kit is the only Shiga toxin EIA validated by the AOAC's Performance Tested Methods Program and the use for STEC detection is limited to pure culture, in contrast to numerous E. coli O157:H7 test kits validated by the same AOAC program or adopted as AOAC Official Methods to be used in a variety of foods (AOAC International, 2012). This scenario is anticipated to change in the near future, given the new regulation and increasing demand for non-O157 STEC testing.
Besides EIAs, a Shiga toxin colony immunoblot is commercially available (Roche Diagnostics GmbH, Mannheim, Germany). This assay has been used for the detection and isolation of STEC colonies from a primary isolation plate (Bettelheim, 2007). The addition of mitomycin C or trimethoprim–sulfamethoxazole was found to stimulate toxin production, thereby facilitating immunoblot detection (Karch et al., 1986; Hull et al., 1993).
Similar to Shiga toxins, E. coli O antigens are common markers for the detection and identification of non-O157 STEC serogroups. Conventional serotyping for O serogroup identification is based on agglutination reactions caused by the antigen–antibody cross-linking (DebRoy et al., 2011a). Commercial latex agglutination kits for certain non-O157 STEC serogroups are now available (Table 1), while complete serotyping is generally conducted at E. coli reference laboratories such as those at the Centers for Disease Control and Prevention or Pennsylvania State University (Atkinson et al., 2006). Latex agglutination kits are rapid and easy to use, in direct contrast to conventional serotyping, which is time consuming and labor intensive. Very recently, USDA scientists described a method to prepare latex agglutination reagents for the top 6 non-O157 STEC serogroups by covalent immobilization of polyclonal immunoglobulin G antibodies onto the polystyrene particles (latex–immunoglobulin G) (Medina et al., 2012). Inclusivity and exclusivity testing showed that all target organisms produced positive results, but three antisera (anti-O26, anti-O103, and anti-O145) cross-reacted with other STEC serogroups.
The immuno-affinity displayed by E. coli O antigens and corresponding antibodies has also been used widely via the IMS process to facilitate the detection and isolation of non-O157 STEC from food. During IMS, microscopic, paramagnetic beads coated with specific antibodies are used to capture target pathogens from food matrices, thereby concentrating the target cells and simultaneously removing inhibitors from complex food samples (Ge and Meng, 2009; Dwivedi and Jaykus, 2011). The recovery efficiency of IMS varies depending on the target pathogen and its antigenic expression, the affinity between pathogen surface antigen and antibody, and the physicochemical properties of the food matrix (Dwivedi and Jaykus, 2011). Commercial IMS products are now available for some non-O157 STEC serogroups (Table 1). Seiken particles (Denka Seiken Co., Ltd., Tokyo, Japan) in combination with selective enrichment and plating were found to be effective for the isolation of O26 and O111 from beef and produce (Hara-Kudo et al., 2000; Catarame et al., 2003; Kanki et al., 2011). The current USDA protocol uses SDIX beads (Newark, DE) coated with antibodies for the top 6 STEC followed by acid treatment and plating on mRBA agar (USDA, 2012a). It is important to note that the number of non-O157 STEC serogroups to which IMS can be applied is limited by the availability of specific and high-affinity antibodies for those serogroups.
Very recently, flow cytometry and enzyme-linked immunosorbent assays (ELISA) have been developed for the rapid detection of the top 6 non-O157 STEC in ground beef using SDIX polyclonal antibodies (Hegde et al., 2012a; Hegde et al., 2012b). The flow cytometry assay was rapid, specific, and quantitative with a sensitivity of 2×103 target cells in pure culture and 1–10 cells in ground beef following 8 h of enrichment (Hegde et al., 2012b). The ELISA assays exhibited 100% specificity for serogroups O103, O111, and O121, 98.2% specificity for serogroups O26 and O45, and 99.1% specificity for serogroup O145. In artificially inoculated ground-beef samples, the detection limits were in the range of 1–10 CFU/25 g following 24-h enrichment (Hegde et al., 2012a). Another study used enzyme-labeled phages (Phazymes) in combination with IMS enrichment on an ELISA platform to detect STEC O26, O103, O111, O145, and O157 (Willford et al., 2012). Depending on the serogroups, the detection accuracy ranged from 75.9% to 93% with detection limits of 105 to 106 CFU/mL in pure culture and 1 CFU/g in spinach samples following 8-h enrichment (Willford et al., 2012).
Molecular-Based Methods
Nucleic acid amplification tests (NAATs) such as PCR, real-time quantitative PCR (qPCR), and isothermal amplification techniques have been adopted to detect non-O157 STEC. The assays target either common virulence genes or O serogroup–specific genes. The most frequently targeted virulence gens are those coding for Shiga toxins (Stx) and intimin. Over the years, many variants of Stx1, Stx2, and intimin have been identified, which currently encompass three Stx1 subtypes (Stx1a, Stx1c, and Stx1d) and seven Stx2 subtypes (Stx2a, Stx2b, Stx2c, Stx2d, Stx2e, Stx2f, and Stx2g) (Feng et al., 2011), as well as 21 intimin subtypes (α1, α2, β1, ξR/β2B, δ/β2O, κ, γ1, γ2, θ, ɛ1, νR/ɛ2, ζ, η1, η2, ι1, μR/ι2, λ, μB, νB, ξB, and o) (Blanco et al., 2006). Other targets used include hemolysin gene (hlyA) and genes on pathogenicity islands (termed “O islands”), such as a number of type III non-LEE-encoded effector (nle) genes (Coombes et al., 2008; Madic et al., 2011).
Since the early 1990s, numerous PCR assays have been developed to detect stx1 and stx2 (Table 2). As the numbers of subtypes increase, the variety and number of PCR assays designed to detect these subtypes also increase (Bettelheim and Beutin, 2003). Although most PCR assays were fast (2–4 h) and sensitive (10–103 CFU/reaction), performances varied when evaluated independently (Bastian et al., 1998; Ziebell et al., 2002). With the advent of qPCR technology in the 2000s, great improvement in the speed and sensitivity of STEC detection has been achieved (Bettelheim and Beutin, 2003). In addition, many of the stx PCR and qPCR assays are amenable to multiplexing with those for eae or other virulence factors, a feature not easily achievable by immunological-based methods. Both FDA and USDA methods use qPCR assays to screen for STEC (Monday et al., 2007; FDA, 2012a; USDA, 2012a). It is of note that when comparing FDA's PCR method and commercially available Shiga toxin EIAs, both types of assays failed to detect some Stx subtypes but not others (Feng et al., 2011). Other studies also reported the failure of some PCR assays to detect genetically distant Stx subtypes (Reischl et al., 2002; Beutin et al., 2009).
N/A, not available; MPN, most probable number; TSB, trypticase soy broth.
As the complete sequences of many (93 to date) E. coli O-antigen gene clusters become available, corresponding NAATs for specific O-serogroups are being developed at a fast pace (DebRoy et al., 2011a). Two genes, wzx and wzy, encoding the O-antigen flippase and the O-antigen polymerase, respectively, were unique across most of the E. coli O-antigen gene clusters, promising good targets for serogroup-specific detection. PCR primers for 58 E. coli O-serogroups targeting these two genes were compiled recently (DebRoy et al., 2011a). Additional genes on the O-antigen gene cluster, such as wbd and wzm, have also been used as targets for assay design (Wang et al., 1998; DebRoy et al., 2011a). Table 2 is a select list of PCR and qPCR assays developed for the top 6 non-O157 STEC serogroups; many are multiplex assays for stx, eae, and other virulence genes.
Recently, a novel NAAT termed loop-mediated isothermal amplification (LAMP) has attracted great attention as a rapid, specific, sensitive, and cost-effective pathogen detection method in food testing and clinical diagnostics (Notomi et al., 2000; Mori and Notomi, 2009). LAMP employs four to six specially designed primers and a strand-displacing Bst DNA polymerase to amplify up to 109 target DNA copies under isothermal conditions (60–65°C) within an hour (Notomi et al., 2000). LAMP assays targeting stx1 and stx2 have been developed and evaluated in food samples (Hara-Kudo et al., 2007; Hara-Kudo et al., 2008; Kouguchi et al., 2010; Zhao et al., 2010). A LAMP STEC detection kit (Eiken Chemical Co., Ltd., Tokyo, Japan) is commercially available (Table 1). Very recently, Wang et al. (2012a; 2012b) developed a suite of LAMP assays by targeting stx1, stx2, eae, and wzx or wzy genes of the top 6 non-O157 STEC serogroups and E. coli O157:H7 and evaluated the assays in ground beef and produce samples. The assays were rapid (40 min), specific, and sensitive (approximately 1–20 CFU per reaction). After 6–8 h of enrichment, the LAMP assays consistently achieved accurate detection of very low levels (1–2 CFU/25 g) of non-O157 STEC cells in ground beef, beef trim, and produce (Wang et al., 2012b).
Despite the advances in developing NAATs for the rapid, specific, and sensitive detection of non-O157 STEC in food, except for those listed in Table 1, the vast majority of methods are not yet commercially available. Other caveats associated with NAATs are the presence of assay inhibitors in food and the inability to differentiate dead from viable cells (Ge and Meng, 2009). The inclusion of internal amplification controls and the addition of viability dyes such as propidium monoazide have been developed to address those concerns (Yaron and Matthews, 2002; Chen et al., 2011; Li and Chen, 2012). Finally, the detection of virulence genes is not a guarantee of gene expression (Feng et al., 2011), and in the case of multiple STEC strains contaminating a single food sample, further confirmation is needed to determine whether the gene profiles detected are from a single strain or multiple different strains (Gould et al., 2009).
On several occasions, NAATs have been combined with immunological methods for the increased sensitivity. For example, a PCR assay (targeting common regions of stx1 and stx2) was coupled with ELISA to develop a highly sensitive and specific method for STEC detection in ground beef (Ge et al., 2002). On the other hand, highly sensitive immuno-PCR assays have been developed to detect Stx2 toxin and its variants based on antibody capture followed by DNA amplification (Zhang et al., 2008; He et al., 2011). Assurance GDS MPX Top 7 STEC (BioControl Systems, Inc., Bellevue, WA) uses IMS first to capture E. coli O157 and the top 6 non-O157 STEC serogroups, which is followed by qPCR assays (Table 1).
Probe hybridization is yet another format of molecular-based detection method. Specifically, colony-lift hybridization procedures have been applied in an effort to increase the isolation of non-O157 STEC strains, particularly when they are present at low levels relative to background flora, as in food samples (Grant et al., 2011). The process is similar to colony immunoblot described above but differs in that probes are used rather than antibodies. Specifically, the samples are screened first by PCR for stx1 and stx2, then positive samples are plated out and hybridized onto membranes with stx-labeled probes (Grant et al., 2011). This approach has been used successfully in multiple studies to facilitate the isolation of non-O157 STEC from various food samples (Begum et al., 1993; Cobbold and Desmarchelier, 2000; Jenkins et al., 2003; Nielsen and Andersen, 2003; Auvray et al., 2007; Stephan et al., 2008; Ju et al., 2012).
Advanced Technologies
Several low-density DNA oligonucleotide microarrays combining the detection of multiple O-serogroups and H types, as well as virulence genes, have been designed for the rapid detection and characterization of STEC strains (Ballmer et al., 2007; Bugarel et al., 2010; Quinones et al., 2012). For example, a GeneDisc array was used to identify 12 O-serogroups (top 6, O55, O91, O104, O113, O118, and O157) and 7 H types (H2, H7, H8, H11, H19, H21, and H28), which are the most clinically relevant STEC serotypes worldwide. Multiple virulence genes were also simultaneously targeted, including stx1, stx2, eae, ehxA (encoding enterohemolysin), and nleB and nleE (located on O island-122), among others (Bugarel et al., 2010). High specificity and concordance with conventional serotyping were reported, suggesting a valuable approach for the identification of STEC strains with a high potential for human disease. It is noteworthy that such assays have not been directly applied in food enrichment broth; rather, they are confirmation tools after pure cultures have been isolated and DNA purification is necessary to obtain reliable results.
Recently, microbead-based suspension arrays using the Luminex technology have been used in an immunoassay to detect Shiga toxins (Clotilde et al., 2011) and in a probe-based assay to detect 10 clinically relevant STEC serogroups (top 6, O91, O113, O128, and O157) (Lin et al., 2011a). The immunoassay recognized Stx1 and several Stx2 subtypes and correctly identified them in 48 samples of ground beef, lettuce, and milk spiked with low levels (<2 CFU/g) of STEC strains (Clotilde et al., 2011). In the probe-based assay (Lin et al., 2011a), PCR primers were first designed to target serogroup-specific wzx or wzy genes. The PCR products were hybridized to probe-labeled bead sets, and fluorescence signals of the beads were detected on a Luminex platform (Bio-Rad Laboratories, Hercules, CA). The assay was rapid (<4 h), specific, robust, and high throughput as demonstrated in a multilaboratory validation study (Lin et al., 2013). As the Luminex technology is designed to detect up to 100 analytes, the array is amenable to expansion with additional O-serogroups of interest (Lin et al., 2011a).
Very recently, polymorphisms of the clustered regularly interspaced short palindromic repeat (CRISPR) regions of STEC strains belonging to serotypes O26:H11, O45:H2, O103:H2, O104:H4, O111:H8, O121:H19, O145:H28, and O157:H7 have been explored as unique markers for assay development (Delannoy et al., 2012a; Delannoy et al., 2012b). DNA sequences of the CRISPR loci of these strains were analyzed and unique regions were selected as targets for designing single qPCR assays for each individual serotype (O:H combination). The assays were shown to be highly specific and sensitive, although cross-activity occurred, particularly to strains of the same H type (Delannoy et al., 2012a; Delannoy et al., 2012b). Further studies are warranted to evaluate the accuracy of these CRISPR-based assays with a large strain set and also their applicability in food testing.
Future Directions
Given the recently implemented USDA regulation on six non-O157 STEC serogroups in raw beef products, testing demand for non-O157 STEC in food is on the upward trend. In response to this growing market, many test-kit-manufacturing companies may rapidly expand their efforts to develop and validate such kits. An ideal detection system for non-O157 STEC should be rapid, specific, sensitive, high throughput, and capable of simultaneously detecting multiple serogroups. Due to the low numbers of non-O157 STEC organisms in food, enrichment may be an indispensable step. One strategy to decrease the sample-to-result time is to design improved media formulations that would facilitate “flash” enrichment. This improved enrichment procedure could then be coupled with rapid screening tools (e.g., Shiga toxin EIA, PCR, LAMP) to obtain definitive negative or presumptive positive results. For presumptive positive samples, further confirmations by traditional culture-based methods are needed. It is therefore important that methods be developed to facilitate the efficient isolation of non-O157 STEC in the presence of high-level background cells by culture. Such methods would include improved IMS, immunoblot, colony hybridization protocols, as well as better-formulated selective and differential media. Any media formulation (or modification) will need to be carefully designed and validated to ensure its applicability across pertinent non-O157 serogroups while still effectively reducing interfering background flora. Also, for the purpose of public health surveillance and disease prevention and control, bacterial cultures remain critical.
It is also desirable that new procedures for detecting non-O157 STEC in food be integrated as much as possible into those already in place for E. coli O157:H7. Although this review focuses on the top 6 non-O157 STEC serogroups in the United States, we acknowledge that non-O157 STEC is a diverse group that is emerging and rapidly evolving. Therefore, detection methods should be easily adaptable to accommodate newly recognized serogroups of significant public health interest, as in the case of E. coli O104:H4, an enteroaggregative E. coli that acquired the stx gene and has been implicated in the recent German (also involving other countries) outbreak (Buchholz et al., 2011).
Technologies routinely used in chemical, physical, and engineering laboratories have been adopted for STEC detection, such as biosensors (Subramanian et al., 2012), mass spectrometry (Pierce et al., 2012), and nanotechnology (Jyoti et al., 2010). Recently, an inexpensive and user-friendly point-of-care testing device (termed Gene-Z) was demonstrated to be able to carry out multiplexed genetic testing using a wireless connection (Stedtfeld et al., 2012). The rapidly evolving next-generation sequencing technology is yet another powerful tool that may potentially transform the methods used for identifying and characterizing foodborne pathogens. Although pure culture is generally required to carry out next-generation sequencing, culture-independent sequencing technologies do exist and have been used to examine the ecology of bacterial population in the environment, food, and the intestinal tracts of animals and humans (Pallen et al., 2010). With collaborative efforts among scientists in multiple disciplines, such promising technologies may be adopted in the near future for the detection of pathogens such as non-O157 STEC in food.
Footnotes
Disclosure Statement
No competing financial interests exist.