
Abstract
To determine whether methicillin-susceptible S. aureus (MSSA) and methicillin-resistant Staphylococcus aureus (MRSA) are present in commercial pig farms and food products from supermarkets in China, we characterized S. aureus isolates from 250 samples associated with swine and animal-related food products in Shandong Province. The isolates were characterized by susceptibility testing, toxin gene detection, pulse-field gel electrophoresis (PFGE), multilocus sequence typing, and spa typing. MRSA were identified and typed by the staphylococcal cassette chromosome mec (SCCmec). The prevalence of S. aureus among all samples was 19.6% (49/250). MRSA and MSSA accounted for 16.7% (20/120) and 8.3% (10/120), respectively, of swine feces samples. Only MSSA was detected from swine carcass, pork, chicken, and raw milk, accounting for 15% (6/40), 10% (3/30), 20% (6/30), and 13.3% (4/30), respectively. The predominant MRSA clone was ST9-t899 SCCmecIVb/PFGE A (70.0%, 14/20). Among the MSSA isolates, ST9-t899/PFGE A was the most prevalent (27.6%), followed by ST15-t084 (17.2%), ST97-t2756 (10.3%), ST1-t127 (6.9%), and ST398-t899 (3.5%). Some lineages were found that are commonly detected in humans (e.g., ST1, ST5, ST7, ST59, ST88) or are human-specific (e.g., ST15). The toxin genes sec, seh, and enterotoxin gene cluster (egc) were significantly more prevalent among isolates of lineage ST9 (p<0.001) compared to other lineages, and the ST9 isolates were more resistant to erythromycin, clindamycin, ciprofloxacin, chloramphenicol, and gentamicin. The same lineage was identified from different sample types, indicating circulation of the related strains within the area of study. In conclusion, swine and food products of animal origin carried S. aureus, and the predominant ST9 clone possesses a multidrug-resistance profile and a high prevalence of sec, seh, and egc enterotoxin genes.
Introduction
To determine whether MSSA and MRSA are present in commercial farms and supermarkets in China, we characterized S. aureus isolates from 250 sample types associated with swine and foods of animal origin. The results of this study contribute to the understanding of the epidemiology of livestock-associated S. aureus and its transmission risk from animals to humans.
Materials and Methods
Collection of samples and isolation of bacterial isolates
Two hundred and fifty nonduplicate samples were obtained from swine feces (February 4, 2012), swine carcasses (November 22, 2011), pork, chicken (June 24, 2011), and milk (October 27, 2010) from two cities in Shandong Province in China. The two cities, Tai'an and Jinan, were less than 100 km apart.
The study protocol was approved by the Medical Ethical Committee of Institute of Animal Science and Veterinary Medicine Shandong Academy of Agricultural Sciences. Collection of animal samples was conducted in accordance with the Chinese Law on Animal Health and Welfare and performed by trained personnel. All the samples in the study were randomly selected. One hundred and twenty fecal samples were obtained per rectum from individual pigs from a commercial farm in Tai'an. They were placed in sterile bags and stored at ambient temperature for transport to the laboratory.
Forty different pig carcasses from a slaughterhouse in Tai'an were also examined. Moreover, 30 randomized samples from raw porcine meat, and 30 raw chicken meat samples were obtained from two supermarkets in Jinan, Shandong. For chicken and pork samples, one piece of meat was examined. The analytical portions from each sample were placed in separate sterile plastic bags, 250 mL of buffered peptone water was added to each bag, and the bags were vigorously shaken. Fifty milliliters of the rinsate from each sample were transferred to separate sterile flasks for isolation and identification of S. aureus.
Thirty cow-milk samples were collected from a bovine farm in Jinan. Each udder quarter was sanitized with 70% isopropyl alcohol. Teat sanitation was accomplished by predipping in 0.5% iodine. The first three streams of milk from a teat to be sampled were discarded to remove contaminated milk from the teat canal, and the subsequent milk sample was collected in a 5-mL sampling vial.
All samples were processed on the day of collection. Bacteria were propagated on selective chromogenic medium used for the detection of S. aureus (bioMérieux, Marcy l'Etoile, France). Presumptive S. aureus colonies were confirmed by colony morphology, Gram staining, catalase production, and coagulase tube test using a VITEK2 automated system (bioMérieux) and by S. aureus-specific polymerase chain reaction (PCR) (Martineau et al., 1998). MRSA isolates were initially identified using cefoxitin (30 μg, Oxoid, Cambridge) and oxacillin disks (1 μg, Oxoid, Cambridge) (CLSI M100-S22, 2012). The presence of the mecA gene was confirmed by PCR as described previously (Perez-Roth et al., 2001).
Antimicrobial susceptibility testing
The antimicrobial susceptibility profiles of S. aureus isolates were determined via the agar dilution method using Mueller–Hinton agar for 18 antimicrobial agents and broth microdilution for tigecycline and daptomycin according to the recommendations and definitions from the Clinical and Laboratory Standards Institute (M100-S22; CLSI M7-A8). The interpretive criteria for the minimum inhibitory concentration of tigecycline were recommended by the U.S. Food and Drug Administration (Raad et al., 2007). The antimicrobial agents tested included oxacillin (Sigma Chemical, St. Louis, MO), cefoxitin (Sigma), cefazolin (Sigma), cefuroxime (Sigma), ceftriaxone (Sigma), erythromycin (Sigma), clindamycin (Sigma), moxifloxacin (Sigma), ciprofloxacin (Sigma), tetracycline (Sigma), chloramphenicol (Sigma), gentamicin (Sigma), trimethoprim/sulfamethoxazole (Sigma), rifampin (Sigma), levofloxacin (Daiichi Pharmaceutical, Tokyo, Japan), tigecycline (Pfizer Pharmaceuticals, New York, NY), linezolid (Pfizer), teicoplanin (Sanofi-Aventis, Schiltigheim, France), and vancomycin (Eli Lilly, Indianapolis, IN). ATCC29213 (S. aureus) and ATCC29212 (Enterococcus faecalis) were used as quality-control organisms.
Molecular typing methods
All S. aureus isolates were characterized by multilocus sequence typing (MLST), spa typing, and pulse-field gel electrophoresis (PFGE). Chromosomal DNA was prepared using the genomic DNA purification kit (Tian Gen, Beijing, China). MLST was performed, as described previously (Enright et al., 2000), to compare the sequences of the PCR products to those at the MLST website (
Detection of toxin genes
All S. aureus isolates were screened for the Panton-Valentine Leukocidin (pvl) gene, staphylococcal enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sej, sep, seq, sem, sen, and seo), and genes for exfoliative toxins (et) and toxic-shock-syndrome toxin (tst). Presence of each gene was determined via PCR and electrophoresis using the primers previously described (Baba et al., 2002; Omoe et al., 2005). All PCR assays were performed in a Mastercycler (Eppendorf, Germany). PCR reagents were purchased from Takara (Dalian, China), and primers were synthesized by Invitrogen (Shanghai, China). The 50-μL PCR mixtures consisted of 5 μL of DNA preparation, 38.1 μL of double-distilled water, 5 μL of 10X buffer containing 2.0 mM MgCl2, 1.0 μL dNTP, 0.25 μL of each primer (100 μM), and 0.3 μL (5 U/μL) of Taq DNA polymerase. After amplification for 30 cycles (30 s of denaturation at 94°C, 30 s of annealing at 55°C, and 1 min of extension at 72°C), the PCR products were resolved by electrophoresis through 1.5% agarose gels (Sigma, Beijing, China).
Statistical analysis
All susceptibility data and molecular test results were analyzed using WHONET, Version 5.6. Pearson chi-square test or Fisher's exact test was used to compare the categorical variables. The significance level α was set at 0.05. The statistical analyses were carried out in SPSS, Version 18.0 (SPSS, Chicago, IL).
Results
Carriage rate of S. aureus among different samples
S. aureus was detected in 19.6% (49/250) of the animal-associated samples. Of the 49 S. aureus isolates obtained from the samples, 20 isolates were confirmed to be MRSA (Table 1). MRSA were only obtained from swine fecal samples and accounted for 16.7%, whereas the prevalence of MSSA among the fecal samples was 8.3%. The MSSA prevalence among the porcine meat from swine carcasses was 15%, whereas the prevalence of MSSA in porcine meat, chicken meat, and raw milk was 10.0% (3/30), 20.0% (6/30), and 13.3% (4/30), respectively.
MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible Staphylococcus aureus.
Resistance profile of S. aureus isolates
All MRSA isolates were resistant to most antibiotics tested, including oxacillin (resistance rate, 95%), cefoxitin (95%), cefazolin (95%), cefuroxime (95%), ceftriaxone (95%), tetracycline (100%), chloramphenicol (90%), erythromycin (90%), clindamycin (95%), gentamicin (80%), and ciprofloxacin (90%). The susceptibility rates of MRSA to moxifloxacin and trimethoprim/sulfamethoxazole were 55% and 95%, respectively. By contrast, all the MSSA isolates were susceptible to most antibiotics tested including oxacillin, cefazolin, ceftriaxone, and cefuroxime (Table 2). MRSA isolates were significantly more resistant to erythromycin, clindamycin, tetracycline, levofloxacin, moxifloxacin, ciprofloxacin, chloramphenicol, and gentamicin than MSSA isolates were (p values <0.001–0.035, Table 2). None of the S. aureus isolates were resistant to rifampin, linezolid, teicoplanin, vancomycin, tigecycline, or daptomycin.
p<0.05 was statistically significant for R%.
Molecular typing of S. aureus
All MRSA were isolated from swine feces from a single farm. The predominant genotype was ST9-t899-SCCmec IVb/PFGE A (70.0%, 14/20), followed by ST9-t7400-SCCmec IVb/PFGE A (10.0%, 2/20), ST9-t034-SCCmec IVb/PFGE A, ST9-t3521-SCCmec IVb/PFGE A, ST9-t5390-SCCmec IVb/PFGE A, and ST5-t899-SCCmec IVb/PFGE A (5.0%, 1/20). Among the LA-MSSA isolates, ST9-t899/PFGE A accounted for 27.6% (8/29), followed by ST15-t084 (17.2%, 5/29), ST97-t2756 (10.3%, 3/29), and ST1-t127 (6.9%, 2/29). A total of 11 PFGE types (A–J) were identified (Table 3). The dominant PFGE type A (A1–A8) represented 28 isolates, of which 27 were from swine feces and one from raw chicken from a supermarket. ST9 MSSA and ST9 MRSA had related PFGE patterns of type A (Fig. 1).
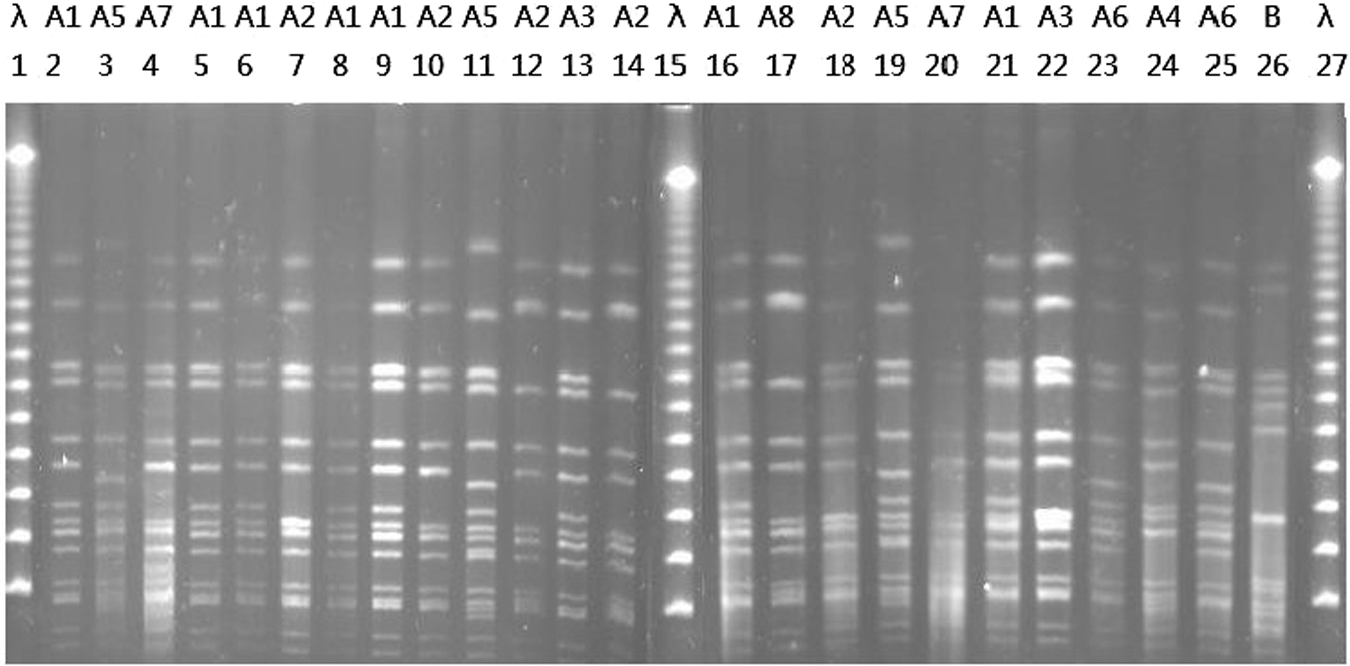
Pulsed-field gel electrophoresis (PFGE) profiles of Sma I-digested chromosomal DNA. Lane λ, lambda ladder PFGE marker (New England Biolabs, Beijing, China); line 2, 3, 5, 6, 7, 8, 9, 10, 11, 12, 14, 16, 18, 19, 21: methicillin-resistant Staphylococcus aureus (MRSA) ST9-t899; line 4, 13, 17, 20, 22, 24, 25: methicillin-susceptible S. aureus ST9-t899; line 23: ST9-t5390 MRSA; line 26: ST5-t899 MRSA.
Toxin genes found in less than five isolates were not listed in this field.
MLST, multilocus sequence typing; PFGE, pulsed-field gel electrophoresis; SCCmec, staphylococcal cassette chromosome mec; N, ceftriaxone; T, tetracycline; F, ciprofloxacin; L, levofloxicin; P, chloramphenicol; E, erythromycin; C, clindamycin; G, gentamicin; H, trimethoprim/sulfamethoxazole; O, oxacillin.
Toxin gene detection
The toxin gene analysis revealed that sec, seh, and enterotoxin gene cluster (egc) genes were prevalent (Table 3), whereas sed, see, sej, and tst were negative in all the isolates. An ST88 isolate harbored sep and pvl, and an ST398 possessed et. The sep toxin gene was found in two ST7 isolates and seq gene in two ST59 isolates, respectively. The positive detection rate of sec, seh, and egc was 51.0%, 57.1%, and 40.8%, respectively. All the MRSA isolates were positive for seh, one was negative for sec, and five were negative for egc, whereas the prevalence of sec, seh, and egc among MSSA was 34.5% (10/29), 58.6% (17/29), and 17.2% (5/29), respectively. The seh toxin gene was highly detected in clonal complex (CC) 9 (55.1%, 27/49), followed by CC15 (6.1%, 3/49), CC1 (4.1%, 2/49), CC5 (4.1%, 2/49), CC7 (4.1%, 2/49), and CC88 (2.1%, 1/49); the seh toxin gene was not detected among isolates of CC59, CC97, and CC398. The sec positive isolates were mostly CC9 (51%, 25/49), followed by CC59 (4.1%, 2/49), CC88 (2.1%, 1/49), and CC5 (2.1%, 1/49). The egc were almost detected in CC9 (36.7%,18/49) but also in two CC5 (4.1%, 2/49).
Comparison of antimicrobial susceptibility and enterotoxin gene prevalence among the predominant clone complexes
Significant differences were found in the susceptibility of CC9 and other clonal complexes to erythromycin, clindamycin, tetracycline, ciprofloxacin, chloramphenicol, and gentamicin (p values, <0.001–0.044, Table 4). CC9 isolates were more resistant to erythromycin, clindamycin, ciprofloxacin, chloramphenicol, and gentamicin, whereas isolates of all other clonal complexes were more resistant to tetracycline. The prevalence of toxin genes varied among the clonal complexes. The prevalence of sec, seh, and egc genes was significantly higher in CC9 than in CC15 or the set of other clonal complexes (p<0.001, Table 4).
Discussion
Data for animal-related food contamination caused by S. aureus is limited in China. We found that the prevalence of S. aureus in pork and milk was lower in our study compared to a study in Shaanxi province, China (Wang et al., 2012a). This may be due to the differences in the geographical distribution of S. aureus. The prevalence of MRSA from swine in China was 11.4% in 2009 (Cui et al., 2009), while our study found it was as high as 16.7% at a single farm.
In our study, the predominant S. aureus genotype was ST9/PFGE A, of which 27 isolates were from swine feces and one from chicken meat. Recently, ST9 isolates were found in samples from pigs, horses, and cows (Pantosti, 2012; Wang et al., 2012b). Documentation of ST9 in chicken meat extends the potential reservoirs and indicates expanded risk to human health. About 90% (18/20) of ST9 MRSA isolates were multidrug resistant, consistent with another study in China (Cui et al., 2009). It has been reported that the pig gut could be a hot-spot for emergence of multidrug-resistant bacteria. The co-localization of multiple resistance genes enables their co-selection and persistence (Lamendella et al., 2011). Livestock-associated ST9 MRSA can act both as a donor and a recipient of antimicrobial resistance genes, and therefore, ST9 MRSA is likely to obtain other resistant genes, resulting in multidrug-resistant ST9 MRSA.
The toxin gene characterization showed that the majority of the isolates carried sec, seh, and egc enterotoxin genes; prior studies did not examine the occurrence of these genes (Cui et al., 2009; Wagenaar et al., 2009; Wang et al., 2012a). We found that the sec, seh, and egc genes were more common in ST9. The sec, seh, and egc were reported to be located on mobile genetic elements: SaPI2, SCCmec, and vSAβ, respectively (Baba et al., 2002; Lindsay et al., 2006; Wang et al., 2009), which may facilitate the mobility of sec, seh, and egc and result in the transmission of these toxin genes among different strains.
The order of evolution for MSSA and MRSA of CC9 is uncertain. CC9-MSSA may have acquired SCCmec, yielding CC9-MRSA, or may have arisen from CC9-MRSA that lost SCCmec or mecA. Hanssen et al. reported the transfer of SCCmec into an MSSA genetic background converting S. aureus to MRSA (Hanssen et al., 2006). In our study, ST9-t899-SCCmec IVb/PFGE A coexisted with ST9-t899/PFGE A MSSA; however, the order of their evolution requires further research. In this study, we did not identify ST9-t899 MRSA from pork or chicken samples, but the limited number of samples from pork and chicken may have resulted in detection of only MSSA strains. MRSA were only detected in pig fecal samples, and the feed was checked and was not the source of MRSA in these samples. All the MRSA isolates were ST9-SCCmecIVb, consistent with a previous study in which the majority of the MRSA from swine carcasses belonged to ST9-t899-SCCmecIVb (Ho et al., 2012b). In contrast, Cui et al. found that all the ST9-t899 MRSA from mainland China harbored SCCmec III (Cui et al., 2009), and Neela et al. found only SCCmec V among pig-associated ST9 isolates in Malaysia (Neela et al., 2009). This variability demonstrates the potential for ST9-MSSA to acquire various SCCmec types through horizontal gene transfer, resulting in ST9- MRSA.
In this study, the presence of the same molecular type in different sample types confirmed that the same S. aureus lineages are able to colonize various sources. For example, ST1-t127/PFGE E MSSA was detected in both pork and chicken from the supermarket. ST7-t189/PFGE F1 MSSA was isolated in chicken, whereas the variants ST7-t10232/PFGE F2 MSSA were detected in pork. MLST types ST1, ST5, ST7, ST15, ST59, and ST88 were associated with pig or chicken samples in our study but were human-associated or reported as human-specific (ST15) in a previous study (Cuny et al., 2010). In a separate study, ST97 was predominantly identified from cattle (Hasman et al., 2010), consistent with our detection of ST97 in milk.
ST9-MSSA can colonize humans and be transmitted between humans and pigs in the United Kingdom (Armand-Lefevre et al., 2005); however, no cases have been reported in Asia. Our lack of samples from farm workers prevented analysis of potential transmission from pigs to humans. Further work should be performed to investigate farm or supermarket workers to assess the transmission of S. aureus between human and animals.
The high prevalence of sec, seh, and egc enterotoxigenic genes among ST9 isolates contrasts with the absence of these genes in swine-associated ST398 isolates (Kock et al., 2009; Schijffelen et al., 2010). Our findings for ST9 also contrast with the reported absence of staphylococcal enterotoxin genes in ST9 isolates from pig nasal samples in Hong Kong (Ho et al., 2012a).
Conclusions
In conclusion, S. aureus occurred in swine and animal-related food products (chicken, pork, and milk), and a multidrug-resistant ST9 clone with sec, seh, and egc enterotoxin genes was predominant. Genetically related ST9 MSSA and ST9 MRSA isolates were detected. Importantly, one ST9/PFGE A MSSA isolate, detected in chicken meat, was genetically related to an ST9 isolate from pig feces, suggesting either broad circulation of related strains or contamination of chicken meat from a pig-associated source.
Footnotes
Acknowledgment
This study was partially supported by the Program for New Century Excellent Talents in University (NCET-10-0205) and Key Projects in the National Science and Technology Pillar Program (2012EP001002).
Author Contributions
H. Wang and Y. Liu conceived and designed the experiments; Y. Liu and J. Qi carried out the collection of strains; W. He, H. Chen, C. Zhao, H. Li, and F. Zhang performed the experiments; W. He, H. Wang, Y. Liu, and J. Qi analyzed the data; H. Wang and W. He wrote the paper.
Disclosure Statement
No competing financial interests exist.