
Abstract
The objective of this study was to clarify the molecular antimicrobial resistance mechanisms of Salmonella enterica serovar Indiana isolated from chickens in China. A total of 327 chicken intestinal content and feces were collected in Shandong, China in 2009. Isolates were serotyped and antimicrobial susceptibility testing was performed. Thirty-five (10.7%) Salmonella isolates were recovered, and 16 (45.7%) were Salmonella enterica serovar Indiana, which were resistant to at least 14 of 15 antimicrobial agents. The 16 Salmonella enterica serovar Indiana detected and other 13 Salmonella enterica serovar Indiana that were selected from 133 Salmonella enterica serovar Indiana isolated in 2008 were subjected to pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). Then class 1 integron and drug resistance genes were detected by polymerase chain reaction. Linkage between plasmids and resistance components was determined by conjugation, electrotransformation, S1 nuclease-PFGE, polymerase chain reaction–based replicon typing and Southern blot assays. Regions flanking integrons were sequenced by modified random primer walking strategy. PFGE and MLST suggested that all the 29 Salmonella enterica serovar Indiana isolates that shared >78% similarity in PFGE patterns were of the same MLST type, ST17. Two kinds of class 1 integrons had different integrase genes and the same variable region (dfrA7/aadA4/IS26/aac(6′)-Ib/bla OXA-1 /catB3/arr-3), and additional antimicrobial resistance genes such as bla CTX-M-24, floR, and so on were detected on IncHI2 plasmids in 29 Salmonella enterica serovar Indiana, and seven plasmids were conjugative. Analysis of the genetic environment of the integrons suggested that these integrons might have been formed by the help of IS26. To our knowledge, the variable region in class 1 integrons is the longest reported in Salmonella to date. The unique integrons and multiple resistance genes co-located on the IncHI2 plasmid contributed to the dissemination of multidrug resistance.
Introduction
It is well established that integrons play a major role in accumulating cassette-borne resistance genes, and class 1 integrons are the most predominant type of integrons among bacteria of the genus Salmonella (Michael et al., 2006). If integrons are located on plasmids, they may play an important role in the dissemination of antimicrobial resistance among Gram-negative bacteria (Cambray et al., 2010).
In a previous work carried out in 2008 (Lu et al. 2011), 133 Salmonella enterica serovar Indiana strains isolates were recovered from chickens sampled in Shandong province of China: All of the isolates exhibited multidrug resistance phenotypes (resistance to at least 10 different antimicrobial agents). The objectives of the present study were (1) to detect the Salmonella enterica serovar Indiana in broiler chickens in other cities in 2009; (2) to compare the genetic relationship with the Salmonella enterica serovar Indiana isolated in 2008, and (3) to clarify the molecular mechanisms that confer multidrug resistance to Salmonella enterica serovar Indiana.
Materials and Methods
Bacterial strains
A total of 327 chicken fecal (n=102) and intestinal content (n=225) samples were collected in 2009 for Salmonella isolation. Samples were derived from intensive chicken farms or abattoirs in five geographically distinct cities (Binzhou, Ji'ning, Laiwu, Zibo, and Taian) in Shandong province of China. The isolation and identification of Salmonella in 2009 were performed as previously reported (Lu et al., 2011). Serotyping of the Salmonella isolates was carried out by slide agglutination with Salmonella antisera (S & A Reagents Lab Ltd., Bangkok, Thailand), and the results were interpreted using the White-Kauffmann-Le Minor scheme (Grimont and Weill, 2007). Moreover, another 13 multidrug-resistant five Salmonella enterica serovar Indiana strains were included to compare the genotypes and further study the antimicrobial resistance mechanisms. These strains were selected from 133 multidrug resistant Salmonella enterica serovar Indiana isolates from chicken meat or feces in Shandong in 2008 (Lu et al., 2011). The 13 Salmonella enterica serovar Indiana isolates represented the dominant and different PFGE clusters; 12 isolates were isolated from an abattoir in Linyi and one isolate was from an abattoir in Yantai.
Antimicrobial susceptibility
Antimicrobial susceptibility testing of Salmonella isolated in 2009 was performed by the broth microdilution method. Minimal inhibitory concentration (MIC) values of tetracycline, ampicillin, amoxicillin/clavulanic acid, cefazolin, ceftiofur, chloramphenicol, florfenicol, nalidixic acid, enofloxacin, rifampicin, streptomycin, gentamicin, amikacin, olaquindox, and trimethoprim/sulfamethoxazole were determined according to the guidelines of Clinical and Laboratory Standards Institute (CLSI 2008a; 2008b). Escherichia coli ATCC 25922 served as quality control strain. As the CLSI did not offer the resistance breakpoint of rifampicin, streptomycin, and olaquindox, isolates with MIC values ≥64 μg/mL were tentatively classified as resistant.
PFGE and multilocus sequence typing (MLST)
Genomic DNA was digested with restriction enzyme XbaI, and then subjected to PFGE according to the standardized PulseNet PFGE protocol (
The MLST was performed using previously described primers for seven housekeeping genes: aroC, dnaN, hemD, hisD, purE, sucA, and thrA (Torpdahl et al., 2005). The data obtained were compared with the Salmonella enterica MLST database (
Polymerase chain reaction (PCR) detection of integrons and resistance-associated genes
All of the 29 Salmonella enterica serovar Indiana were subjected to PCR assays to amplify the class 1 integrase gene (intI1), the variable region (5′CS-3′CS), and the qacEΔ1/sul1 genes in the 3′ conserved sequence (3′CS) of class 1 integrons as described (Antunes et al., 2006). For those isolates negative for intI1, newly designed primers: ΔintI1-F (5′-TGGACCAGTTGCGTGAGC-3′) and ΔintI1-R (5′-TCAAGGGCGTCGGGAAG-3′) were used to amplify the class 1 integrase gene. Other resistance genes encoding tetracycline resistance tet(A), florfenicol resistance gene floR, β-lactamases (bla CTX-M, bla SHV, bla TEM), olaquindox– chloramphenicol–quinolone resistance gene oqxA-oqxB were detected for all the isolates using previously described primers (Aarestrup et al., 2003; Jouini et al., 2007; Zhao et al., 2010), and gentamicin resistance gene aac(3)-IVa was detected using newly designed primers: aac(3)-IVa-F(5′-GCTCATCGGTCAGCTTCTCA-3′) and aac(3)-IVa-R(5′-AGGGCTGTCGCCACAATGT-3′).
Plasmid characterization and location of resistance components
Transfer of resistance was tested by filter mating assays. The Escherichia coli J53AzR (resistant to azide) was used as recipient strain and each of the 29 Salmonella enterica serovar Indiana isolates served as donors. In addition, DNA of the nonconjugative plasmids from isolates S414 in 2008 and TA155 in 2009 was extracted using QIAGEN Plasmid Maxi Kit (Qiagen, Hilden, Germany) and then electrotransformed into E. coli DH10B. Transconjugants and transformants were selected on MacConkey agar containing 100 mg/L sodium azide, 20 mg/L ampicillin, and 16 mg/L chloramphenicol. The plasmids in the donor strains, transconjugants, and transformants were analyzed by PCR-based replicon typing (Carattoli et al., 2005). The sizes of the plasmids were determined using PFGE of S1 nuclease digests of total DNA (S1-PFGE) (Barton et al., 1995). MIC values of the transconjugants and transformants were determined, and the presence of the integrons and antimicrobial resistance genes was detected as described above. Southern blot hybridization was performed using a 2.5-kb aac-arr3 probe including aac(6′)-Ib/bla OXA-1 /catB3/arr3, which was obtained from PCR amplicons by primers aac-F(5′-GAGCGATCCGATGCTACGAG-3′) and arr3-R(5′-TCTAACGCCGCCATAAACG-3′) after the S1-PFGE.
Molecular analysis of the genetic environment of integron
According to PFGE and plasmid profiles, the transconjugant (J53-pS471) of donor strain S471, and the transformants (DH10B-pS414 and DH10B-pSTA155) of strains S414 and TA155 were chosen for sequence analysis of the regions flanking the integrons. This was done by a modified random primer walking strategy started from intI1 and sul1 using primers sen1 (5′-TCCGCTATTGGTCTCGGTGTC-3′), sen2 (5′-ATCGACGAGGTTGTGCGGCTGAT-3′), ant1 (5′-ATCAGGACCACTACCGATTACG-3′), ant2 (5′-AAGTGGTTCGCATCCTCGGT-3′), and the nested random primers arb1 (5′-GGCCACGCGTCGACTAGTACNNNNNNNNNNGATAT-3′) and arb2 (5′-GGCCACGCGTCGACTAGTAC-3′) as previously described (Zhang et al., 2009). The accuracy of the obtained sequence was confirmed by long-range PCR using the primer pairs located inside or outside of the integron. Sequences of integrons in this study were deposited under GenBank accession number JX885645 and KC237285.
Results
Isolation and antimicrobial susceptibility
In 2009, we recovered a total of 35 (10.7%) Salmonella isolates from chickens in five cities in Shandong. Among the 35 isolates, 16 (45.7%) were Salmonella enterica serovar Indiana, and others were Salmonella enterica serovar Enteritidis. Interestingly, the 19 Salmonella enterica serovar Enteritidis isolates demonstrated resistance to only ampicillin and nalidixic acid, but were susceptible to the other 13 antimicrobials tested (data not shown). On the contrary, multidrug resistance patterns were observed in all 16 Salmonella enterica serovar Indiana isolates (Table 1). With the exception of two isolates from 2009 that were susceptible to tetracycline, the Salmonella enterica serovar Indiana isolates exhibited high MIC values to each of the 15 antimicrobials tested (Table 2).
Minimum inhibitory concentrations of STR, RIF, and OLQ are ≥64 mg/L.
Conjugative.
Nonconjugative.
PFGE, pulsed-field gel electrophoresis; TET, tetracycline; AMP, ampicillin; AMC, amoxicillin/clavulanic acid, CFZ, cefazolin; FUR, ceftiofur; CHL, chloramphenicol; FFC, florfenicol; NAL, nalidixic acid, ENO, enofloxacin; RIF, rifampicin, STR, streptomycin; GEN, gentamicin; AMK, amikacin; OLQ, olaquindox; SXT, trimethoprim/sulfamethoxazole.
Except three isolates with the MIC value of 4 μg/mL.
Except for one conjugant with the MIC value of 4 μg/mL, the MIC value of corresponding donor strains is 4 μg/mL.
No resistance breakpoint has been defined by the Clinical and Laboratory Standards Institute.
MIC, minimum inhibitory concentration; TET, tetracycline; AMP, ampicillin; AMC, amoxicillin/clavulanic acid, CFZ, cefazolin; FUR, ceftiofur; CHL, chloramphenicol; FFC, florfenicol; NAL, nalidixic acid, ENO, enofloxacin; RIF, rifampicin, STR, streptomycin; GEN, gentamicin; AMK, amikacin; OLQ, olaquindox; SXT, trimethoprim/sulfamethoxazole.
PFGE and MLST
The PFGE patterns of the isolates in different years showed some variation, but the similarity of PFGE profiles among the 29 Salmonella enterica serovar Indiana from chickens in China in 2008–2009 was above 78% (Fig. 1). Notably, 14 Salmonella enterica serovar Indiana isolates from four different cities in 2009 shared the same fragment pattern I. MLST indicated all the 29 Salmonella enterica serovar Indiana isolates belonged to a single MLST type, sequence type ST17.
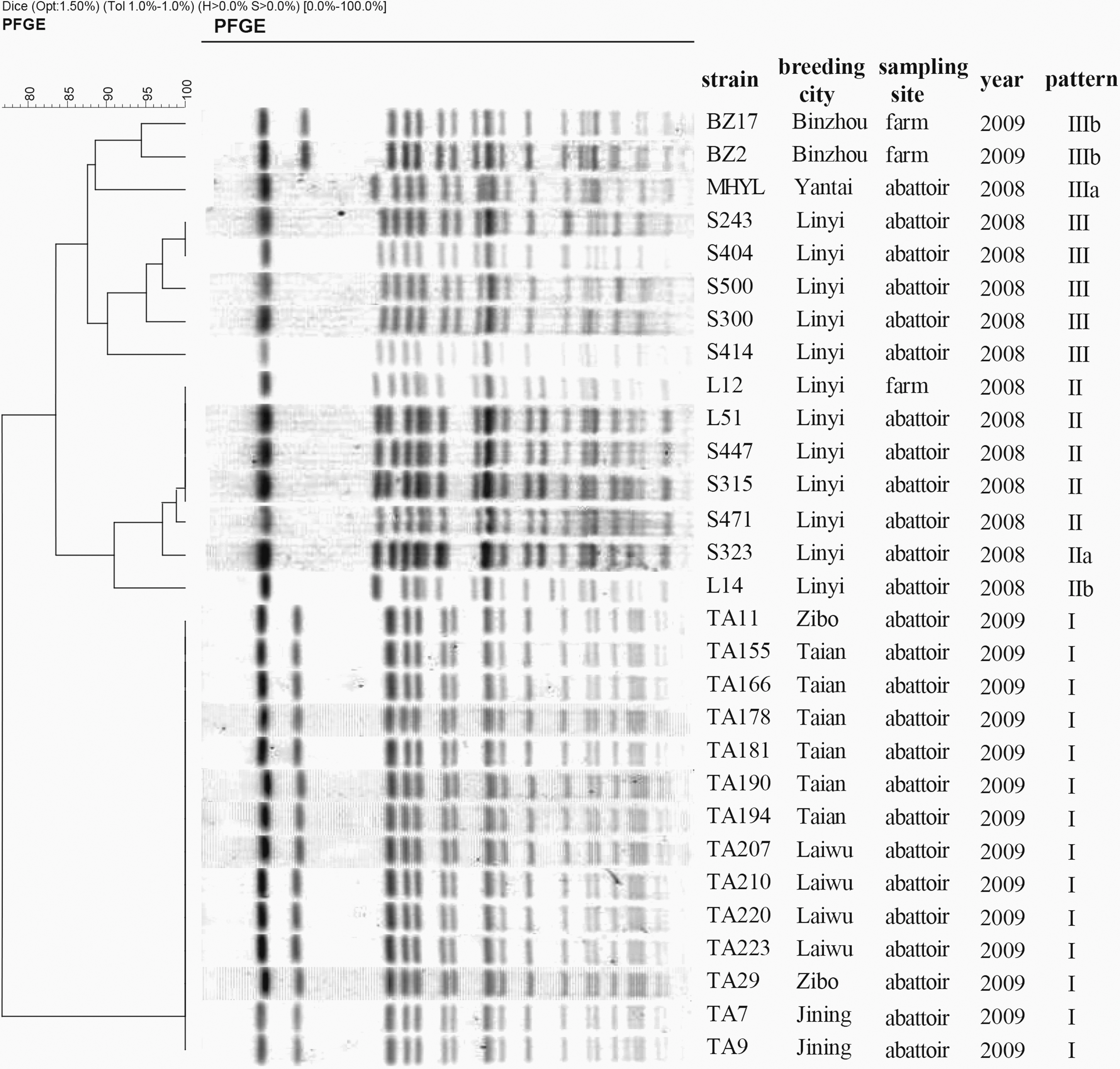
Comparison of XbaI–pulsed-field gel electrophoresis (PFGE) patterns of 29 Salmonella Indiana isolates from chicken feces, intestinal contents, or meat in Shandong Province of China in 2008–2009.
The presence of integrons and resistance genes
PCR screening of integron components in the 29 Salmonella enterica serovar Indiana isolates indicated that 5′CS-3′CS and 3′CS were positive. However, there were some differences of intI1 in isolates from 2008 and isolates from 2009. When using previously described primers (Antunes et al., 2006), the intI1 in 13 isolates in 2008 were positive (amplicons are 898 bp), while the intI1 in 16 isolates in 2009 were negative. However, using the primers of ΔintI1-F and ΔintI1-R, amplicons of 601 bp can be obtained in all 29 isolates. That suggested the intI1 in 2009 might be a variant class 1 integrase gene. Sequence analysis revealed the variable regions in class 1 integrons of all the 29 isolates were the same 5.7 kb and contained six gene cassettes and an IS26 element: dfrA7/aadA4/IS26/aac(6′)-Ib/bla OXA-1/catB3/arr-3. In addition, another six resistance genes—bla TEM-1, bla CTX-M-24, oqxA, oqxB, floR, and aac(3)-IVa—were also identified in each of the 29 Salmonella enterica serovar Indiana isolates. The gene tet(A) was observed in isolates with tetracycline resistance phenotype (n=27) (Table 1).
Plasmid characterization, resistance genes, and integrons analysis
Seven transconjugants from donor strains (L12, L14, L51, S315, S471, S447, S323) were successfully transferred by conjugation, while plasmids in other Salmonella enterica serovar Indiana (n=22) were nonconjugative in our experiments. S1-PFGE analysis showed each of the seven transconjugants harbored one ∼250-kb plasmid and the corresponding seven donor isolates harbored two plasmids (one is ∼250 kb, another is ∼160 kb) per strain. PCR-based replicon typing revealed the plasmids in transconjugants were IncHI2 type. The seven donor Salmonella enterica serovar Indiana isolates each carried an IncHI2 plasmid and another IncA/C plasmid. Two E. coli DH10B electrotransformants from S414 and TA155 were obtained. Plasmids in transformants of S414 and TA155 belonged to IncHI2 type, and the sizes were ∼250 kb and 240 kb, respectively. The original 22 Salmonella enterica serovar Indiana also harbored only one ∼250 kb (n=8) or 240 kb (n=14) IncHI2 plasmid (Fig. 2). Southern blot hybridization further confirmed the variable region of the class 1 integron located on each IncHI2 plasmid (Fig. 2). All other six (bla TEM-1, bla CTX-M-24, oqxA, oqxB, floR, aac(3)-IVa) or seven resistance genes (tet(A), bla TEM-1, bla CTX-M-24, oqxA, oqxB, floR, aac(3)-IVa) identified in the donor strains were detected in transconjugants and transformants. Compared with J53AzR and DH10B, transconjugants and transformants exhibited at least fourfold higher MIC values for all 14 antimicrobial agents tested (Table 2).
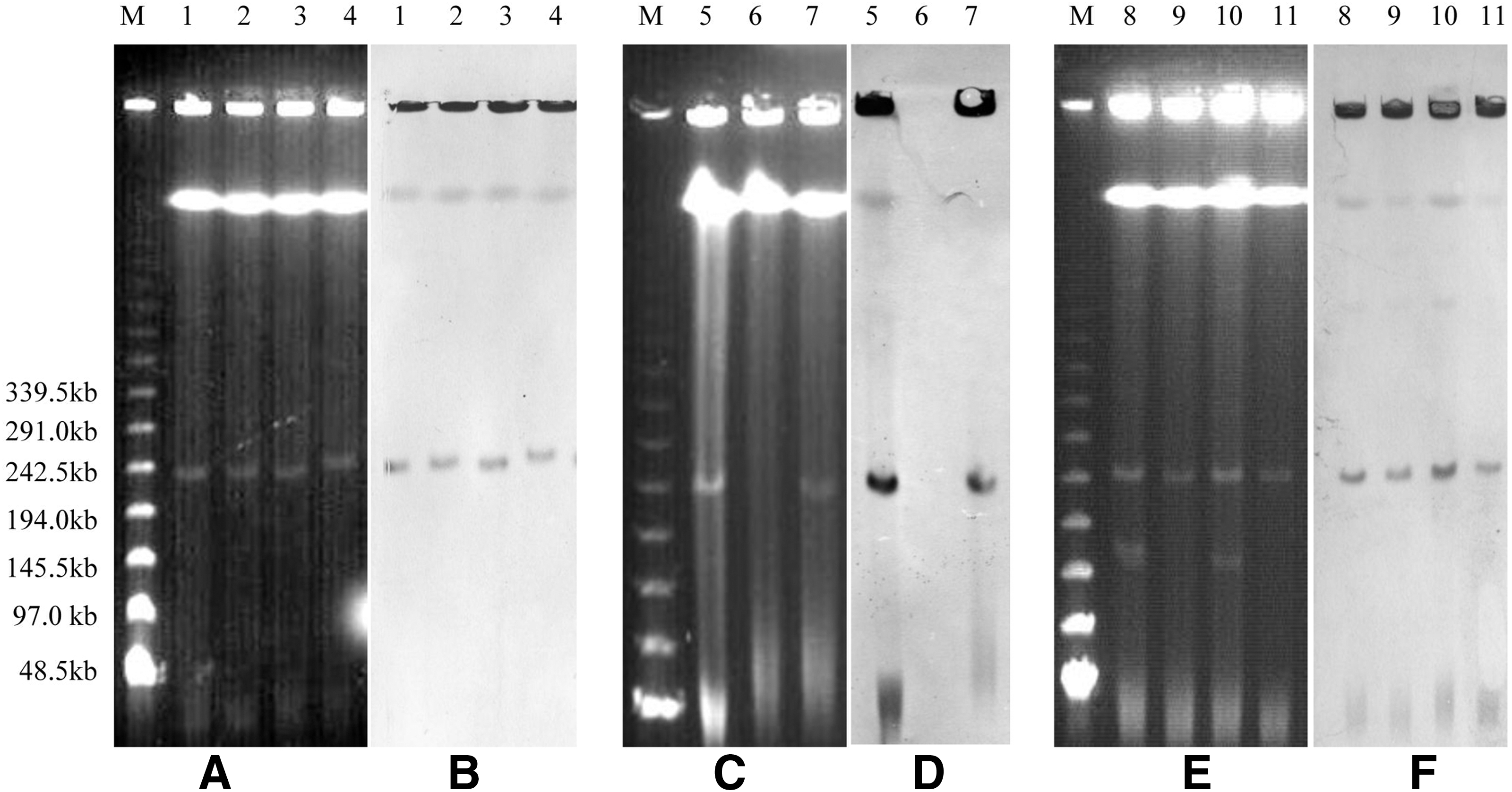
Partial S1–pulsed-field gel electrophoresis (PFGE) and Southern blot profiles of Salmonella enterica serovar Indiana isolates, Escherichia coli transconjugants, and transformants. (
According to modified random primer walking strategy and long-range PCR, sequence of the whole integron, upstream of intI1 and downstream of sul1 were obtained, and also explained why the intI1 in 2009 were negative for the 898 bp amplicon. A 7839 bp (from intI1 to sul1) class 1 integron including intI1 (1014 bp) in S471 and S414, and 7524 bp class 1 integron including a variant intI1 (699 bp, ΔintI1) in TA155 were identified. The orf5 downstream of sul1 in Salmonella enterica serovar Indiana in 2008 and 2009 was truncated (204 bp). We noted that those two kinds of class 1 integron were both flanked by two IS26 elements, but the direction of two IS26 in Salmonella enterica serovar Indiana isolated in different year was adverse (Fig. 3).
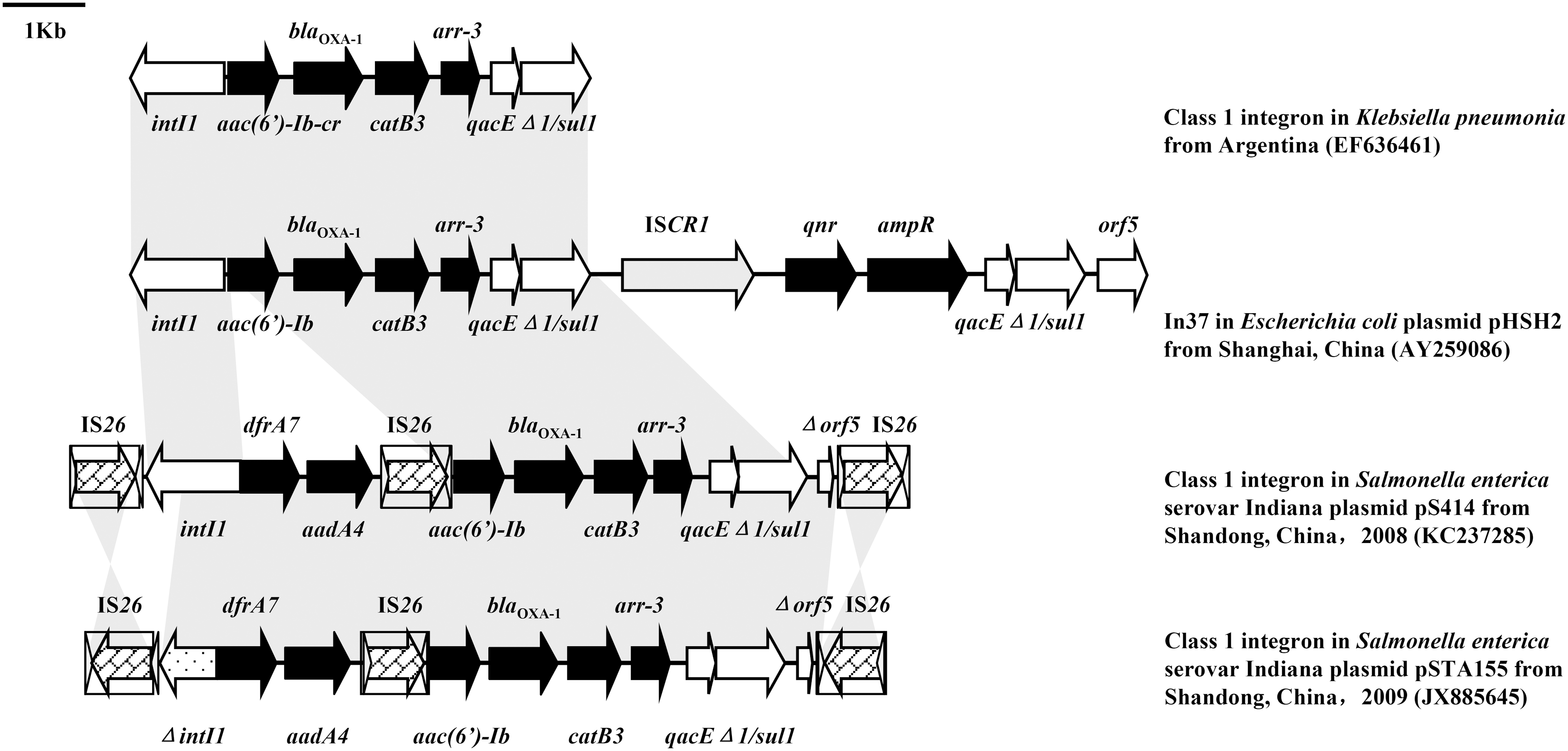
Genetic environment of the integrons in plasmid DNA of Salmonella enterica serovar Indiana S414 and TA155, and the structure comparison with similar class 1 integrons or resistance. The arrows indicate the positions and directions of transcription of the genes. Gene cassettes are displayed in black shading. IS26 elements are shown as white boxes with shading arrows indicating the transposase genes and short white arrowhead indicating inverted repeats of IS elements. The regions of 98% homology are marked by gray shading. A distance scale in kb is indicated above the map.
Discussion
The increase in the occurrence of multidrug-resistant Salmonella enterica serovar Indiana in China stimulated us to study the molecular mechanism of multidrug resistance of Salmonella enterica serovar Indiana in the Shandong province, which is the most important area for chicken production in China. The present study gives a better understanding of the multidrug resistance mechanism of Salmonella enterica serovar Indiana.
Salmonella enterica serovar Indiana once caused food poisoning outbreaks in America and Europe (Price and Carter, 1967; Beckers et al., 1985). The first Salmonella enterica serovar Indiana isolates in China were identified in 2005 in food of animal origin (Yan et al., 2010); among them, three isolates were resistant to multiple antimicrobial agents. Then isolation of Salmonella enterica serovar Indiana in chicken increased in Shanxi province, and Salmonella enterica serovar Indiana showed the highest rate of antimicrobial resistance and multidrug resistance compared to other serotypes (Yang et al., 2010). Xia et al. (2009) reported 12 Salmonella enterica serovar Indiana that were isolated from diarrhea patients in hospitals in Henan province from 2006 to 2007. In 2008, our laboratory isolated a large number of Salmonella enterica serovar Indiana (133 isolates) in Shandong province; and all the isolates were resistant to multiple antimicrobial agents. In the present study, the isolation rate of Salmonella enterica serovar Indiana and Salmonella serovar Enteritidis in 2009 was in accordance with the result of the study in our laboratory in 2008 (Lu et al., 2011), while higher than other reports (Yang et al., 2010). The Salmonella enterica serovar Indiana still showed a much more expanded resistance phenotype compared with Salmonella serovar Enteritidis. Though the PFGE of Salmonella enterica serovar Indiana showed a little variation in different years and cities, MLST suggested those isolates were closely related. This was in accordance with the work of Harbottle et al. (2006), who reported that MLST defined fewer types than PFGE, while MLST may be more relevant to investigations exploring evolutionary and population biology relationships. The ST17 type Salmonella enterica serovar Indiana presumably was the dominant multidrug resistant Salmonella disseminated in chicken farms in Shandong province of China.
We found two kinds of unusual class 1 integrons on IncHI2 plasmids of Salmonella enterica serovar Indiana isolated from 2008 and 2009. The difference of integrons in Salmonella enterica serovar Indiana of 2008 and 2009 was the intI1 gene. The 1014 bp intI1 gene in Salmonella enterica serovar Indiana isolates in 2008 (n=13) was normal. The 699-bp integrase gene in isolates in 2009 (n=16), which has never reported in Salmonella before, exhibited 100% nucleotide identity with the intI1 in E. coli (accession no. EF150886) (Perichon et al., 2007). The intI1 variant still preserved the promoter of itself (Pint) and gene cassettes (Pc). The 7839-kb and 7524-kb class 1 integron located on the IncHI2 plasmids carry gene cassettes that contained the resistance genes dfrA7, aadA4, aac(6′)-Ib, bla OXA-1, catB3 and arr-3, and the sul1 gene in the 3′CS segment and mediated resistance to trimethoprim, streptomycin/spectinomycin, amikacin, ampicillin, chloramphenicol, rifampicin, and sulfonamides, respectively. The previously reported class 1 integrons in Salmonella commonly contain one or two antibiotic resistance cassettes. To the best of our knowledge, the variable region in these two class 1 integrons is the longest variable region, and mediates the most expanded drug-resistance phenotype in Salmonella to date.
The gene cassettes array of aac(6′)-Ib/bla OXA-1 /catB3/arr-3 (4229 bp) exhibited >99.0% nucleotide sequence identity to that of the variable region of a clinical Klebsiella pneumoniae isolate from Argentina (accession no. EF636461) (Quiroga et al., 2007), and to In37 from a clinical E. coli from China (accession no. AY259086) (Wang et al., 2003) (Fig. 3). Taken together, these results suggested this complex Salmonella integron might have been formed with the help of IS26, and at least some of its components probably originated from other species of Enterobacteriaceae such as Klebsiella or Escherichia.
Furthermore, the presence of genes bla TEM-1, bla CTX-M-24, floR, aac(3)-IVa, oqxA-oqxB, and tet(A) in IncHI2 plasmid extended the resistance phenotype to cephalosporins, florfenicol, gentamicin, olaquindox, quinolones, and tetracyclines. Those multiple resistance genes and the unique integron co-located on the IncHI2 plasmid contributed to the multidrug resistance phenotype of Salmonella enterica serovar Indiana. Plasmids that carry such integrons and the additional resistance genes and hence confer such an expanded multiresistance phenotype have, to the best of our knowledge, not been reported in Salmonella before.
A number of Salmonella enterica serovar Indiana with expanded multiresistance phenotype isolated in chicken in Shandong might be associated with the wide use of antimicrobial agents in chicken farms. In our investigated areas, ampicillin, florfenicol, sulfanilamides, and streptomycin had been widely used to prevent or treat chicken disease for a long time. Under the selection pressure of antimicrobials, the multiresistant IncHI2 plasmid can persist in Salmonella enterica serovar Indiana. As an important foodborne pathogen, Salmonella enterica serovar Indiana has been implicated in food poisoning and causes sporadic disorders such as gastroenteritis and miscarriage in humans (Campbell and Eckman, 1975), and can be transmitted from food-producing animals to humans through the food chain. Therefore, the presence of multidrug-resistant Salmonella enterica serovar Indiana will seriously compromise therapeutic options. Moreover, since seven of 29 IncHI2 plasmids were conjugative, a transfer of these multiresistance plasmids from Salmonella enterica serovar Indiana to other bacteria cannot be excluded. Once bacterial pathogens that carry these multiresistance plasmids infect people, antibiotic treatment failures might happen. Thus, continuous monitoring of the dissemination of this multidrug-resistant Salmonella enterica serovar Indiana ST17 and its multiresistance IncHI2 plasmid is warranted.
Footnotes
Acknowledgments
The authors are grateful to Prof. Yuqing Liu (Shandong Academy of Agricultural Sciences, Ji'nan, P. R. China) for helping with sample collection, and Prof. Minggui Wang (Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai, China) and Prof. G.A. Jacoby for kindly providing the reference strain E. coli J53AzR. This work was supported by National Natural Science Foundation of China (30972216), National Science and Technology Pillar Program (2012BAK01B02), and Exclusive Research Foundation for Public Welfare from Ministry of Agriculture of People's Republic of China (201203040).
Disclosure Statement
No competing financial interests exist.